

Phosphorylation of Bcl-associated death protein (Bad) by erythropoietin-activated c-Jun N-terminal protein kinase 1 contributes to survival of erythropoietin-dependent cells
- PMID: 21095239
- PMCID: PMC3039111
- DOI: 10.1016/j.biocel.2010.11.011
Phosphorylation of Bcl-associated death protein (Bad) by erythropoietin-activated c-Jun N-terminal protein kinase 1 contributes to survival of erythropoietin-dependent cells
Abstract
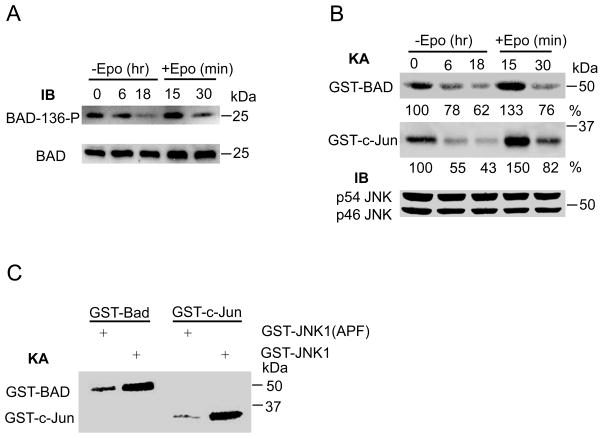
The glycoprotein erythropoietin (Epo) is a hematopoietic cytokine necessary for the survival of erythrocytes from immature erythroid cells. The mitogen-activated c-Jun N-terminal kinase 1 (JNK1) plays an important role in the proliferation and survival of erythroid cells in response to Epo. However, the precise mechanism of JNK1 activation promoting erythroid cell survival is incompletely understood. Here, we reported that JNK1 is required for Epo-mediated cell survival through phosphorylation and inactivation of the pro-apoptotic, Bcl-2 homology domain 3 (BH3)-only Bcl-associated death protein (Bad). Upon Epo withdrawal, HCD57 cells, a murine Epo-dependent cell line, displayed increased apoptotic cell death that was associated with decreased JNK1 activity. Epo withdrawal-induced apoptosis was promoted by inhibition of JNK1 activity but suppressed by expression of a constitutively active JNK1. Furthermore, Epo-activated JNK1 phosphorylated Bad at threonine 201, thereby inhibiting the association of Bad with the anti-apoptotic molecule B-cell lymphoma-extra large (Bcl-X(L)). Replacement of threonine 201 by alanine in Bad promoted Epo withdrawal-induced apoptosis. Thus, our results provide a molecular mechanism by which JNK1 contributes to the survival of erythroid cells.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures



Similar articles
-
JNK and p38 are activated by erythropoietin (EPO) but are not induced in apoptosis following EPO withdrawal in EPO-dependent HCD57 cells.Blood. 2000 Aug 1;96(3):933-40. Blood. 2000. PMID: 10910907
-
Erythropoietin-induced phosphorylation/degradation of BIM contributes to survival of erythroid cells.Exp Hematol. 2009 Feb;37(2):151-8. doi: 10.1016/j.exphem.2008.10.008. Epub 2008 Dec 18. Exp Hematol. 2009. PMID: 19100675 Free PMC article.
-
Jun N-terminal kinase promotes proliferation of immature erythroid cells and erythropoietin-dependent cell lines.Blood. 2004 Aug 1;104(3):696-703. doi: 10.1182/blood-2003-05-1754. Epub 2004 Apr 1. Blood. 2004. PMID: 15059850
-
Unraveling distinct intracellular signals that promote survival and proliferation: study of erythropoietin, stem cell factor, and constitutive signaling in leukemic cells.J Hematother Stem Cell Res. 2000 Feb;9(1):21-9. doi: 10.1089/152581600319586. J Hematother Stem Cell Res. 2000. PMID: 10738968 Review.
-
The role of tyrosine phosphorylation in proliferation and maturation of erythroid progenitor cells--signals emanating from the erythropoietin receptor.Eur J Biochem. 1997 Nov 1;249(3):637-47. doi: 10.1111/j.1432-1033.1997.t01-1-00637.x. Eur J Biochem. 1997. PMID: 9395308 Review.
Cited by
-
Endogenous erythropoietin signaling facilitates skeletal muscle repair and recovery following pharmacologically induced damage.FASEB J. 2012 Jul;26(7):2847-58. doi: 10.1096/fj.11-196618. Epub 2012 Apr 9. FASEB J. 2012. PMID: 22490927 Free PMC article.
-
The Role of c‑Jun Signaling in Cytidine Analog-Induced Cell Death in Melanoma.ACS Omega. 2025 Jul 17;10(29):31776-31788. doi: 10.1021/acsomega.5c02807. eCollection 2025 Jul 29. ACS Omega. 2025. PMID: 40757321 Free PMC article.
-
IMB-6G, a novel N-substituted sophoridinic acid derivative, induces endoplasmic reticulum stress-mediated apoptosis via activation of IRE1α and PERK signaling.Oncotarget. 2016 Apr 26;7(17):23860-73. doi: 10.18632/oncotarget.8184. Oncotarget. 2016. PMID: 27009865 Free PMC article.
-
A Novel Platinum Resistance-Related Immune Gene Signature for Overall Survival Prediction in Patients with Ovarian Cancer.Biochem Genet. 2024 Feb;62(1):112-124. doi: 10.1007/s10528-023-10379-9. Epub 2023 Jun 4. Biochem Genet. 2024. PMID: 37270714 Free PMC article.
-
Differential proteomic analysis of human erythroblasts undergoing apoptosis induced by epo-withdrawal.PLoS One. 2012;7(6):e38356. doi: 10.1371/journal.pone.0038356. Epub 2012 Jun 18. PLoS One. 2012. PMID: 22723854 Free PMC article.
References
-
- Acs G, Acs P, Beckwith SM, Pitts RL, Clements E, Wong K, Verma A. Erythropoietin and erythropoietin receptor expression in human cancer. Cancer Res. 2001;61:3561–5. - PubMed
-
- Cheng EH, Wei MC, Weiler S, Flavell RA, Mak TW, Lindsten T, Korsmeyer SJ. BCL-2, BCL-X(L) sequester BH3 domain-only molecules preventing BAX- and BAK-mediated mitochondrial apoptosis. Mol Cell. 2001;8:705–11. - PubMed
-
- Cieslak D, Lazou A. Regulation of BAD protein by PKA, PKCdelta and phosphatases in adult rat cardiac myocytes subjected to oxidative stress. Mol Cells. 2007;24:224–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
