

Pervasive and cooperative deadenylation of 3'UTRs by embryonic microRNA families
- PMID: 21095586
- PMCID: PMC3698950
- DOI: 10.1016/j.molcel.2010.11.003
Pervasive and cooperative deadenylation of 3'UTRs by embryonic microRNA families
Abstract
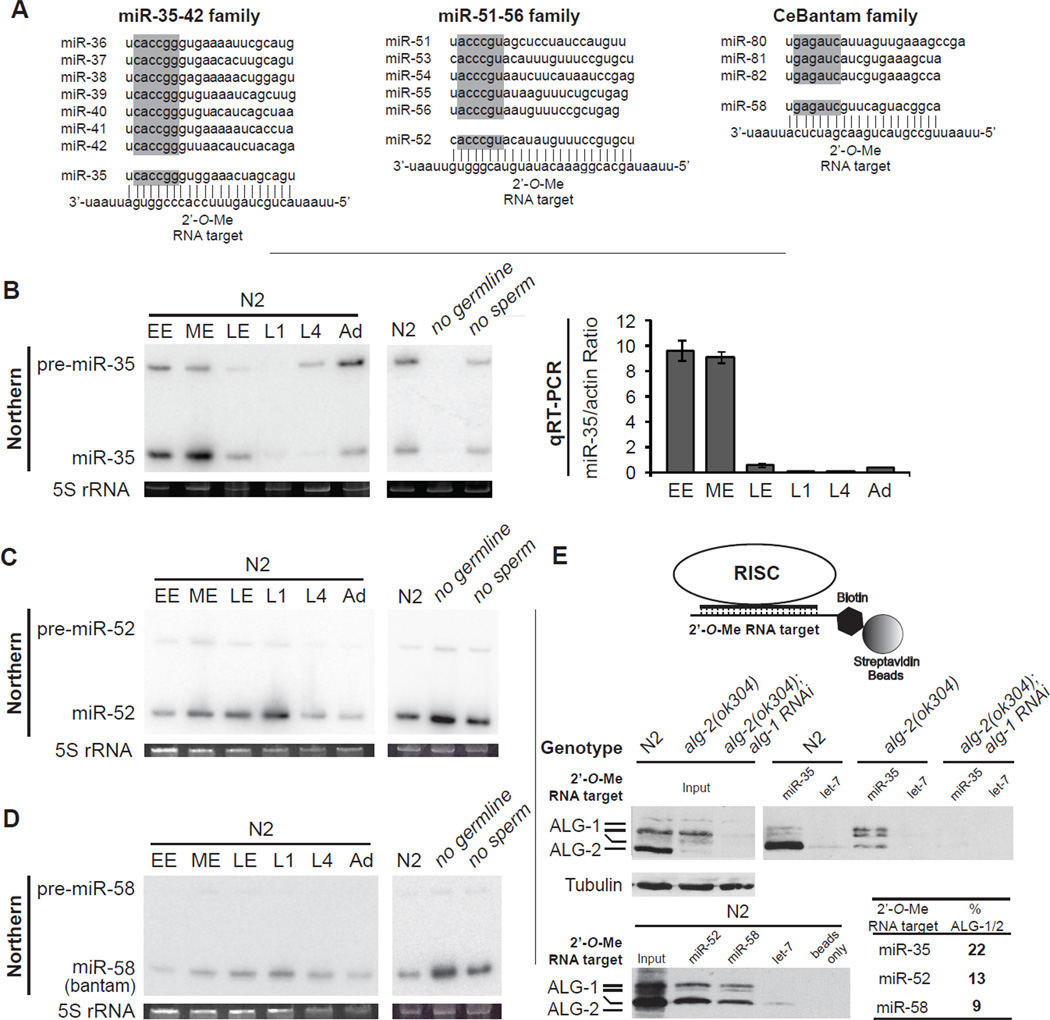
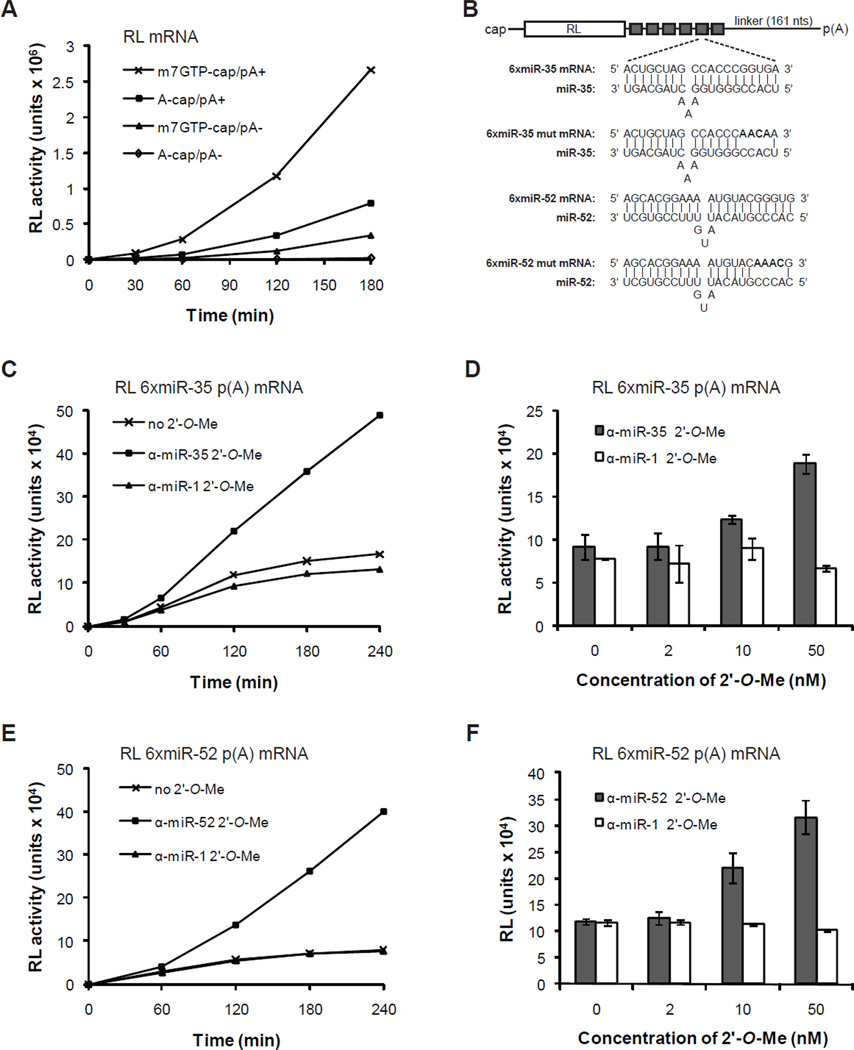
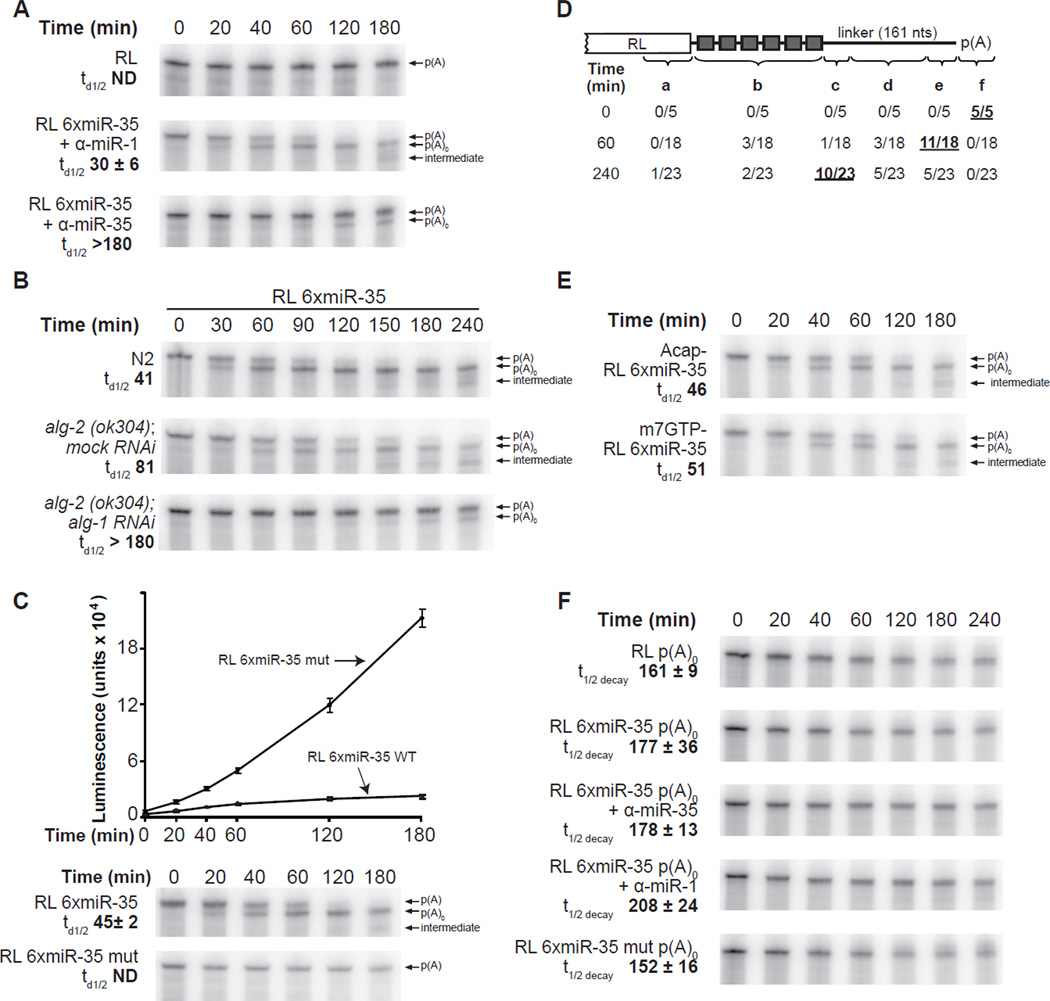
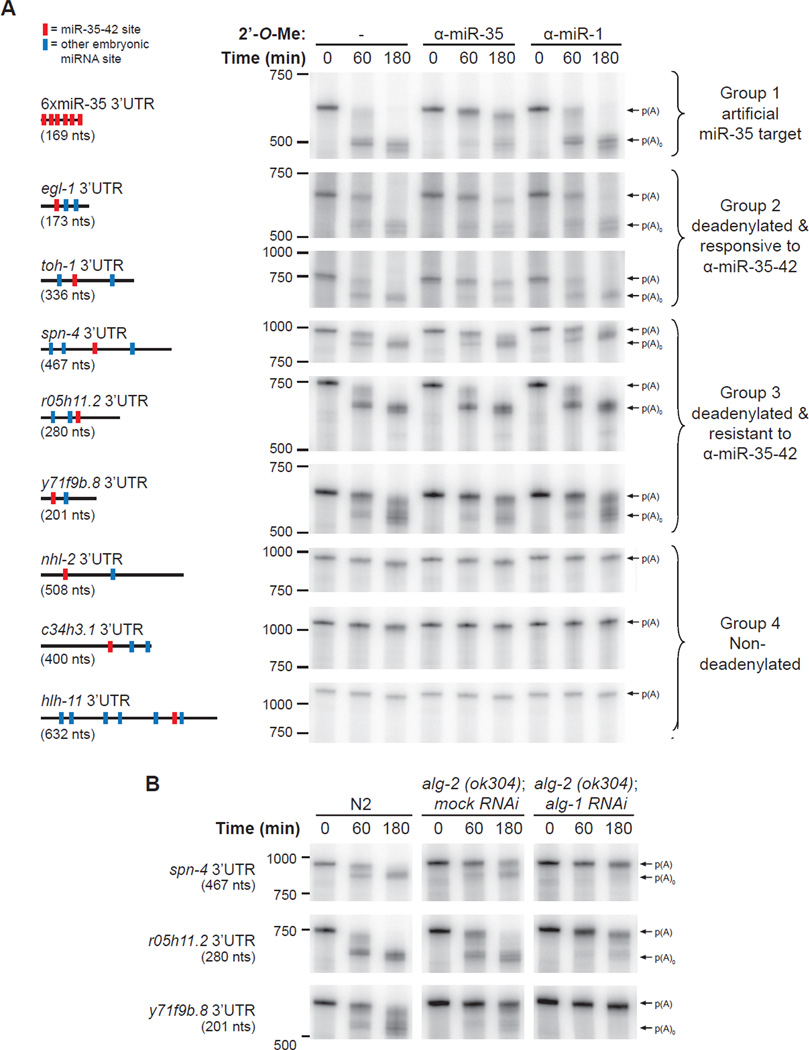
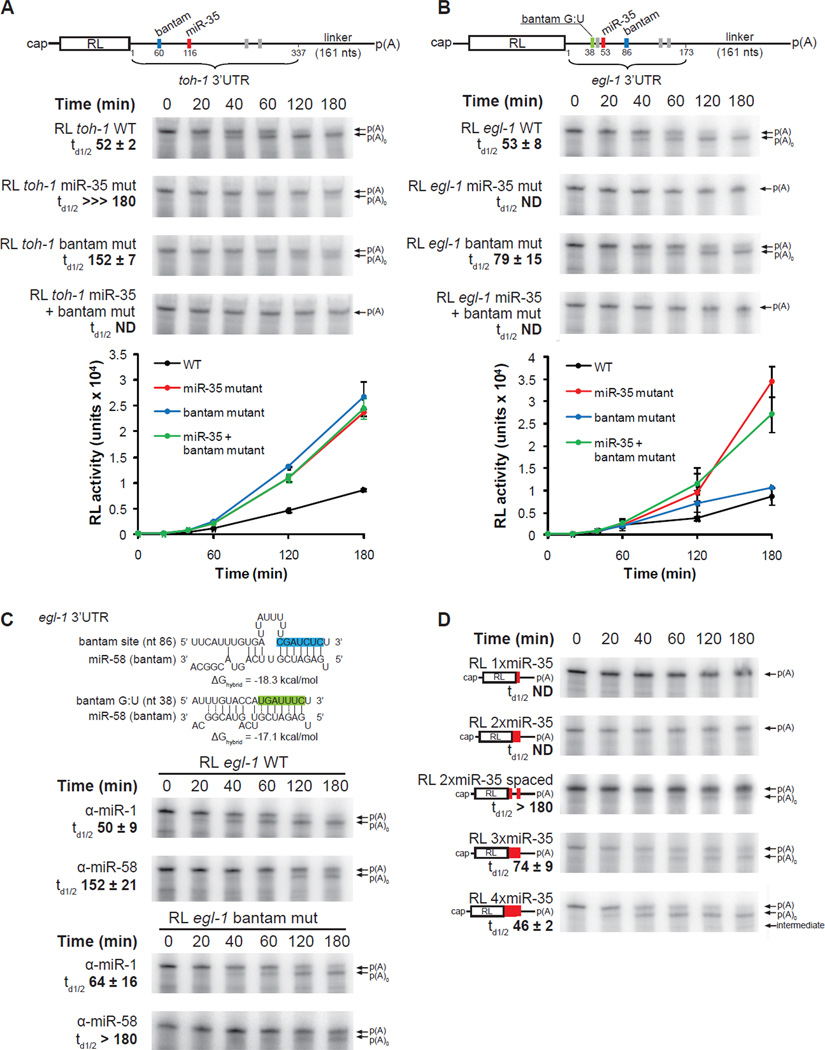
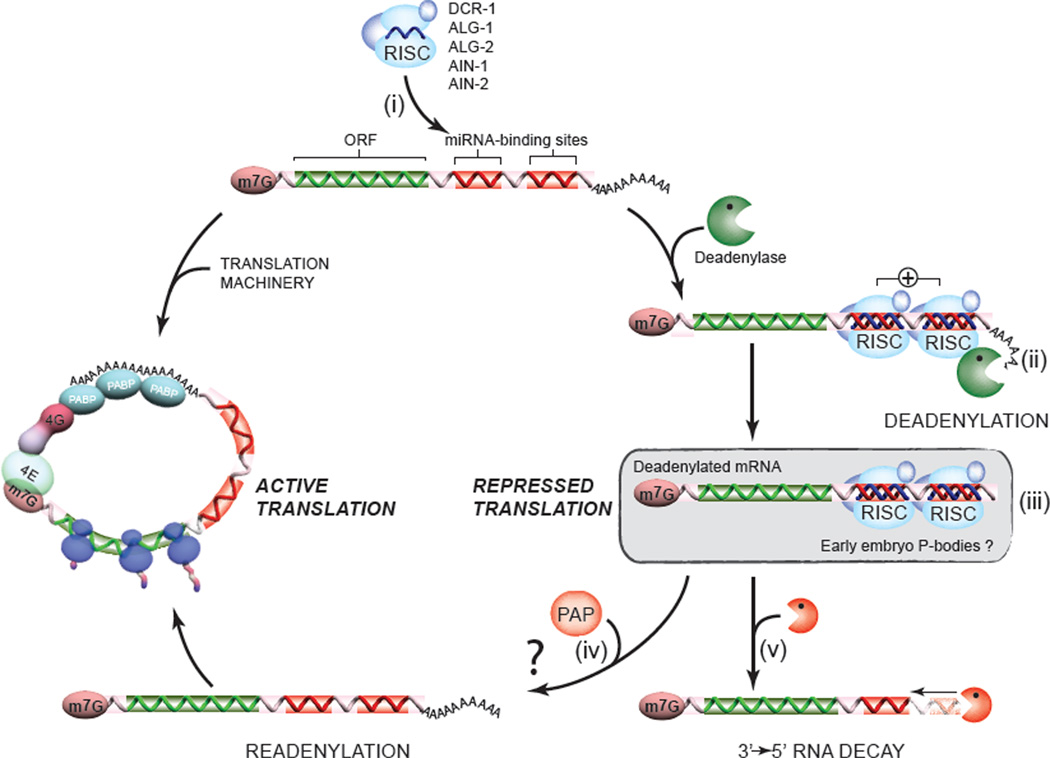
To understand how miRNA-mediated silencing impacts on embryonic mRNAs, we conducted a functional survey of abundant maternal and zygotic miRNA families in the C. elegans embryo. We show that the miR-35-42 and the miR-51-56 miRNA families define maternal and zygotic miRNA-induced silencing complexes (miRISCs), respectively, that share a large number of components. Using a cell-free C. elegans embryonic extract, we demonstrate that the miRISC directs the rapid deadenylation of reporter mRNAs with natural 3'UTRs. The deadenylated targets are translationally suppressed and remarkably stable. Sampling of the predicted miR-35-42 targets reveals that roughly half are deadenylated in a miRNA-dependent manner, but with each target displaying a distinct efficiency and pattern of deadenylation. Finally, we demonstrate that functional cooperation between distinct miRISCs within 3'UTRs is required to potentiate deadenylation. With this report, we reveal the extensive and direct impact of miRNA-mediated deadenylation on embryonic mRNAs.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Ambros V, Lee RC, Lavanway A, Williams PT, Jewell D. MicroRNAs and Other Tiny Endogenous RNAs in C. elegans. Curr Biol. 2003;13:807–818. - PubMed
-
- Bagga S, Bracht J, Hunter S, Massirer K, Holtz J, Eachus R, Pasquinelli AE. Regulation by let-7 and lin-4 miRNAs results in target mRNA degradation. Cell. 2005;122:553–563. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
