

Splicing-dependent RNA polymerase pausing in yeast
- PMID: 21095588
- PMCID: PMC3000496
- DOI: 10.1016/j.molcel.2010.11.005
Splicing-dependent RNA polymerase pausing in yeast
Abstract
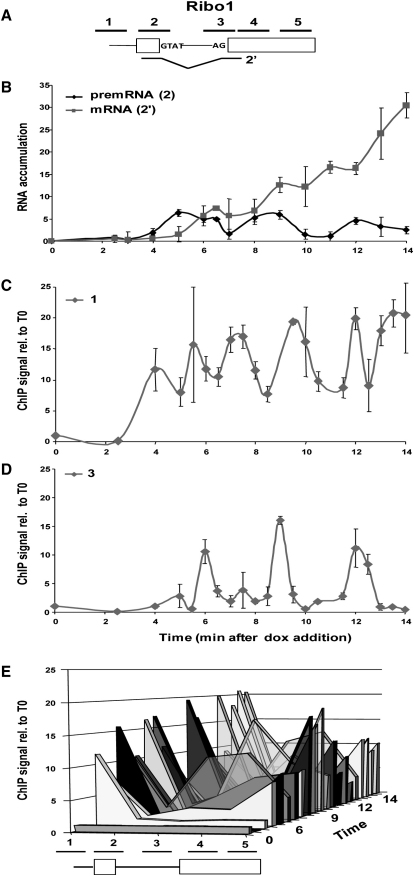
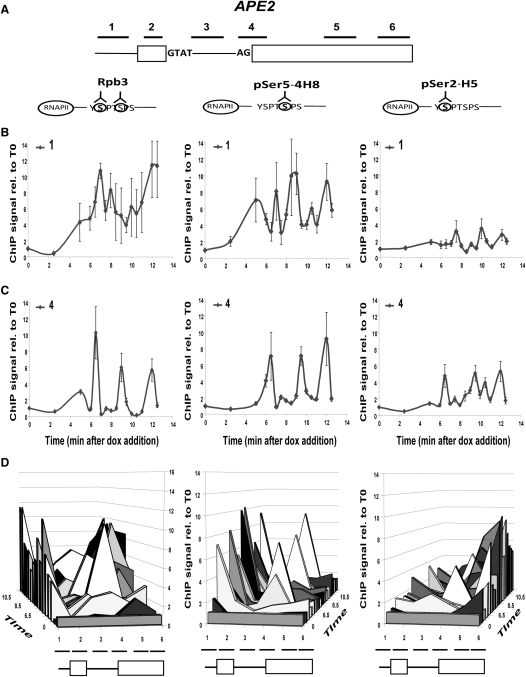
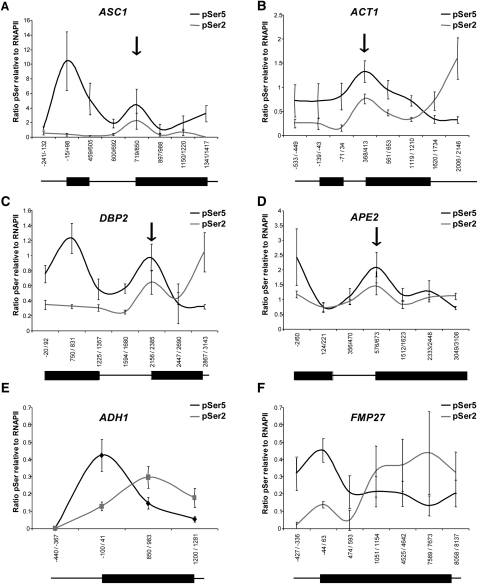
In eukaryotic cells, there is evidence for functional coupling between transcription and processing of pre-mRNAs. To better understand this coupling, we performed a high-resolution kinetic analysis of transcription and splicing in budding yeast. This revealed that shortly after induction of transcription, RNA polymerase accumulates transiently around the 3' end of the intron on two reporter genes. This apparent transcriptional pause coincides with splicing factor recruitment and with the first detection of spliced mRNA and is repeated periodically thereafter. Pausing requires productive splicing, as it is lost upon mutation of the intron and restored by suppressing the splicing defect. The carboxy-terminal domain of the paused polymerase large subunit is hyperphosphorylated on serine 5, and phosphorylation of serine 2 is first detected here. Phosphorylated polymerase also accumulates around the 3' splice sites of constitutively expressed, endogenous yeast genes. We propose that transcriptional pausing is imposed by a checkpoint associated with cotranscriptional splicing.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
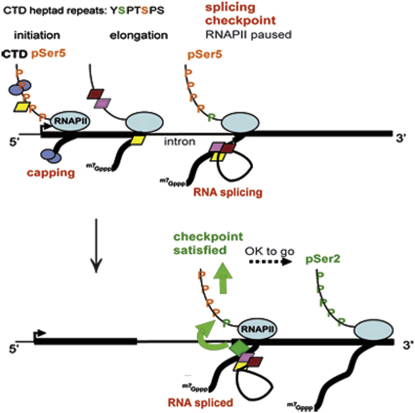

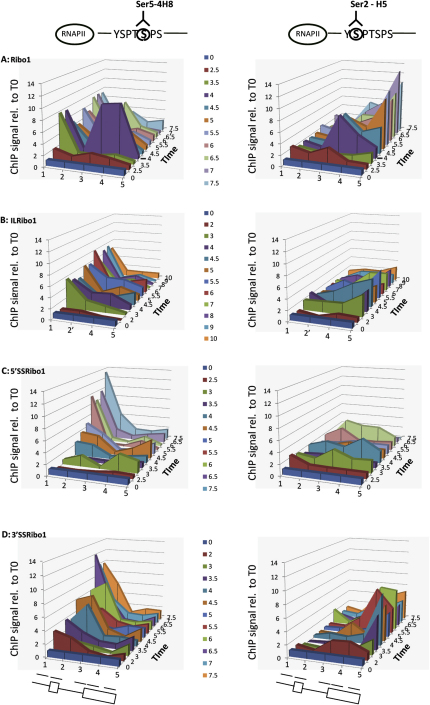
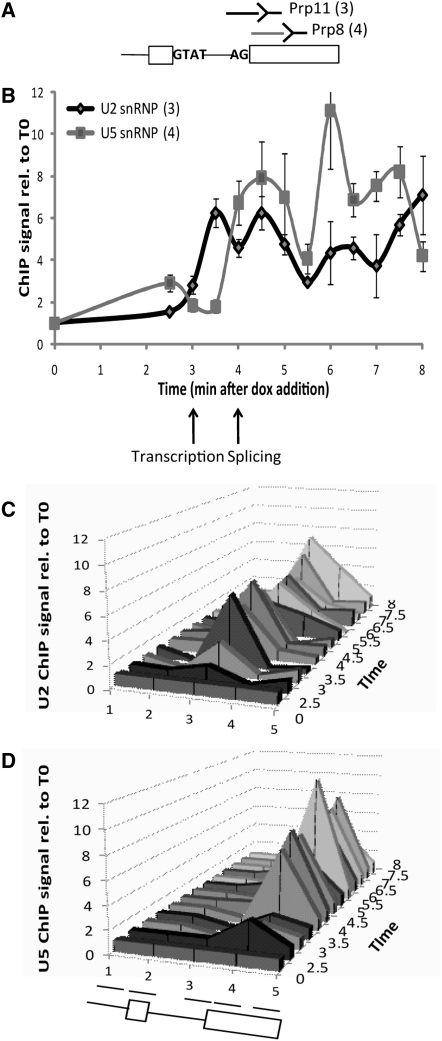
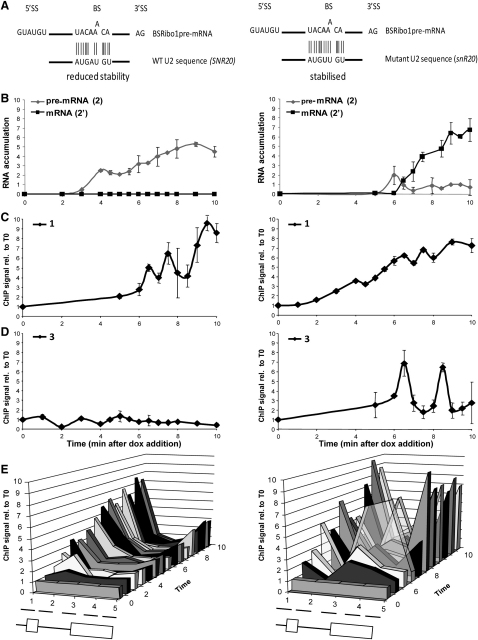
Comment in
-
A pause to splice.Mol Cell. 2010 Nov 24;40(4):503-5. doi: 10.1016/j.molcel.2010.11.019. Mol Cell. 2010. PMID: 21095579
References
-
- Alexander R.D., Barrass J.D., Dichtl B., Kos M., Obtulowicz T., Robert M.C., Koper M., Karkusiewicz I., Mariconti L., Tollervey D. RiboSys, a high-resolution, quantitative approach to measure the in vivo kinetics of pre-mRNA splicing and 3′-end processing in Saccharomyces cerevisiae. RNA. 2010 in press. Published online October 25, 2010. - PMC - PubMed
-
- Batsché E., Yaniv M., Muchardt C. The human SWI/SNF subunit Brm is a regulator of alternative splicing. Nat. Struct. Mol. Biol. 2006;13:22–29. - PubMed
-
- Beyer A.L., Osheim Y.N. Splice site selection, rate of splicing, and alternative splicing on nascent transcripts. Genes Dev. 1988;2:754–765. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
