

ABI4 mediates abscisic acid and cytokinin inhibition of lateral root formation by reducing polar auxin transport in Arabidopsis
- PMID: 21097710
- PMCID: PMC3015119
- DOI: 10.1105/tpc.110.074641
ABI4 mediates abscisic acid and cytokinin inhibition of lateral root formation by reducing polar auxin transport in Arabidopsis
Abstract
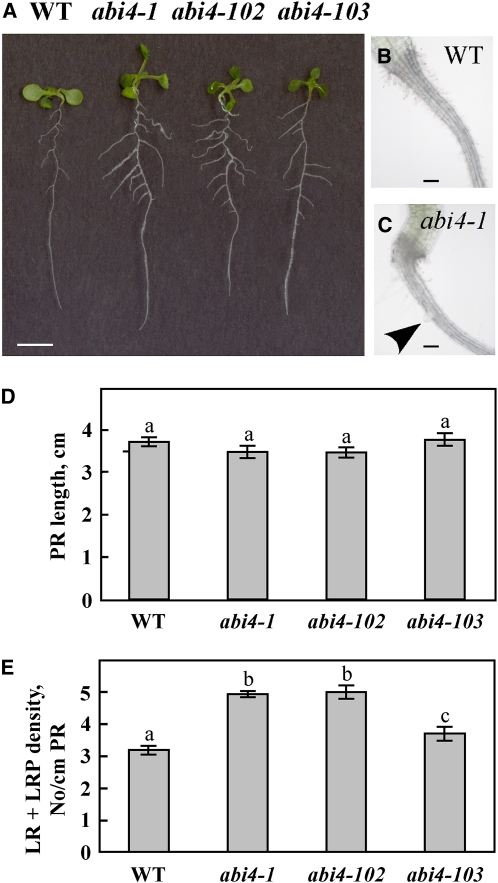
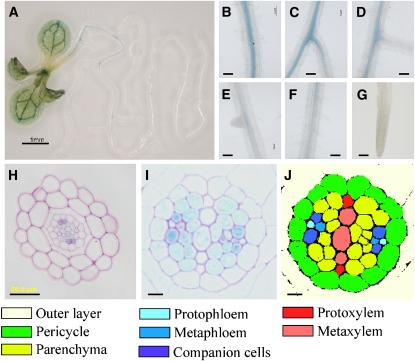
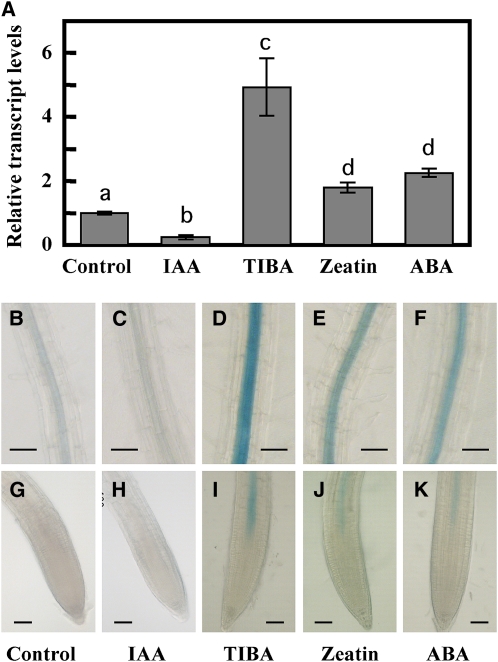
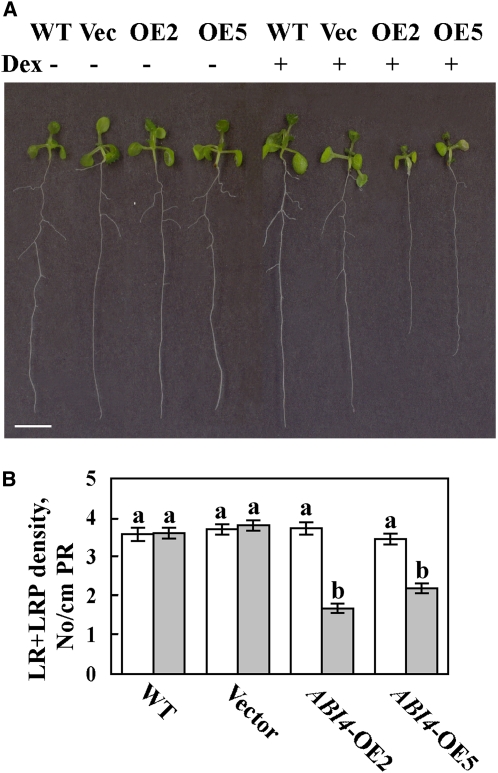
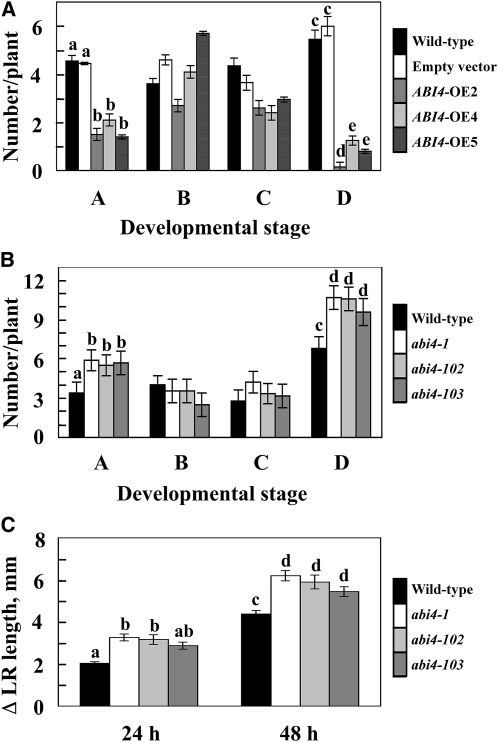
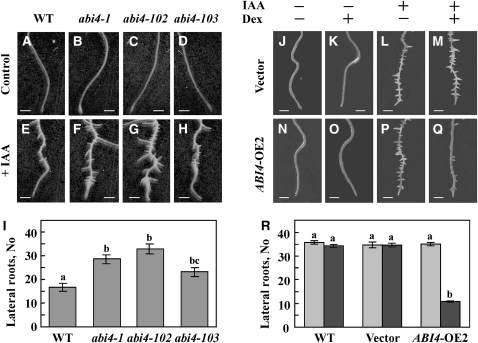
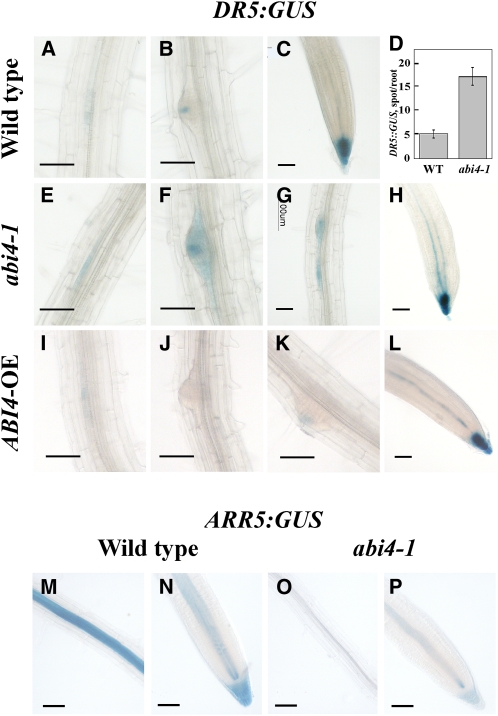
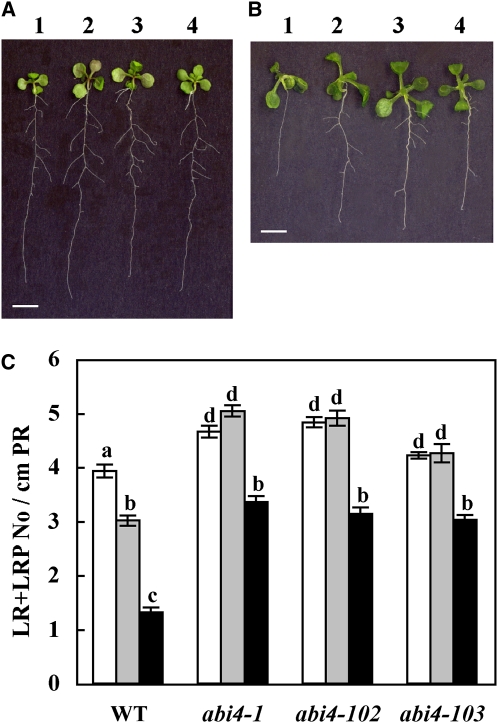
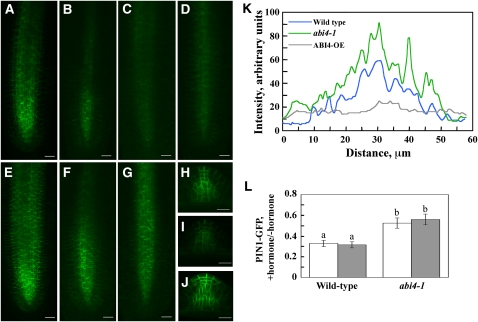
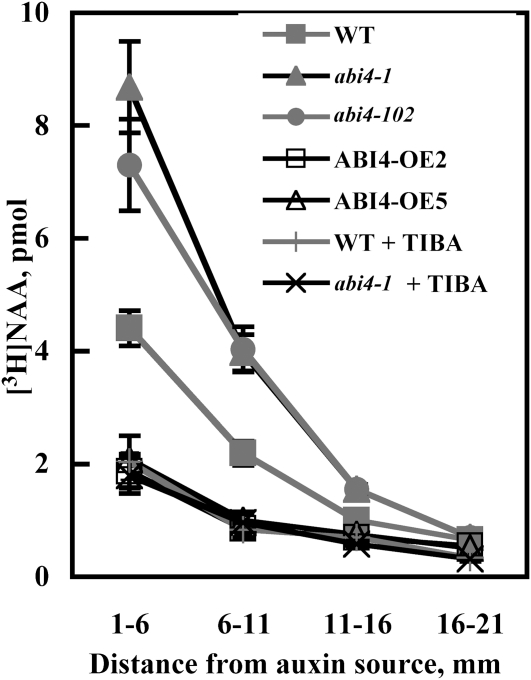
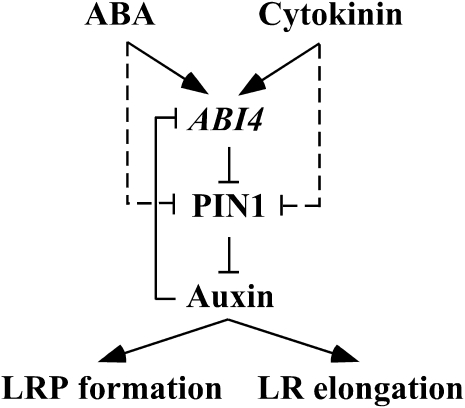
Key steps in a plant's development and adaptation to the environment are the initiation and development of lateral roots (LRs). LR development is regulated by auxin, the major plant hormone promoting LR formation, its counteracting hormones cytokinin, and abscisic acid (ABA). Here, we show that mutating ABSCISIC ACID INSENSITIVE4 (ABI4), which encodes an ABA-regulated AP2 domain transcription factor, results in an increased number of LRs. We show that ABI4 is expressed in roots and that its overexpression impairs LR development. Root expression of ABI4 is enhanced by ABA, and cytokinin and is repressed by auxin. Using hormone response promoters, we show that ABI4 also affects auxin and cytokinin profiles in the root. Furthermore, LR development in abi4 mutants is not altered or inhibited by cytokinin or ABA. Expression of the auxin-efflux carrier protein PIN1 is reduced in ABI4 overexpressors, enhanced in abi4 mutants, and is less sensitive to inhibition by cytokinin and ABA in abi4 mutants than in wild-type plants. Transport levels of exogenously applied auxin were elevated in abi4 mutants and reduced in ABI4 overexpressors. We therefore suggest that ABI4 mediates ABA and cytokinin inhibition of LR formation via reduction of polar auxin transport and that the resulting decrease in root auxin leads to a reduction in LR development.
Figures
References
-
- Abas L., Benjamins R., Malenica N., Paciorek T., Wiśniewska J., Wirniewska J., Moulinier-Anzola J.C., Sieberer T., Friml J., Luschnig C. (2006). Intracellular trafficking and proteolysis of the Arabidopsis auxin-efflux facilitator PIN2 are involved in root gravitropism. Nat. Cell Biol. 8: 249–256 Erratum. Nat. Cell Biol. 8: 249–256 - PubMed
-
- Beeckman T., Viane R. (2000). Embedding thin plant specimens for oriented sectioning. Biotech. Histochem. 75: 23–26 - PubMed
-
- Benková E., Hejátko J. (2009). Hormone interactions at the root apical meristem. Plant Mol. Biol. 69: 383–396 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
