

Contrasting dynamics of radial O2-loss barrier induction and aerenchyma formation in rice roots of two lengths
- PMID: 21097947
- PMCID: PMC3002478
- DOI: 10.1093/aob/mcq221
Contrasting dynamics of radial O2-loss barrier induction and aerenchyma formation in rice roots of two lengths
Abstract
Background and aims: Many wetland species form aerenchyma and a barrier to radial O(2) loss (ROL) in roots. These features enhance internal O(2) diffusion to the root apex. Barrier formation in rice is induced by growth in stagnant solution, but knowledge of the dynamics of barrier induction and early anatomical changes was lacking.
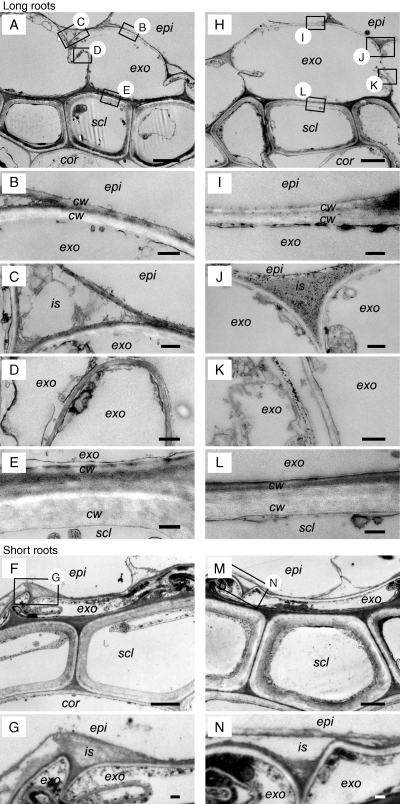
Methods: ROL barrier induction in short and long roots of rice (Oryza sativa L. 'Nipponbare') was assessed using cylindrical root-sleeving O(2) electrodes and methylene blue indicator dye for O(2) leakage. Aerenchyma formation was also monitored in root cross-sections. Microstructure of hypodermal/exodermal layers was observed by transmission electron microscopy (TEM).
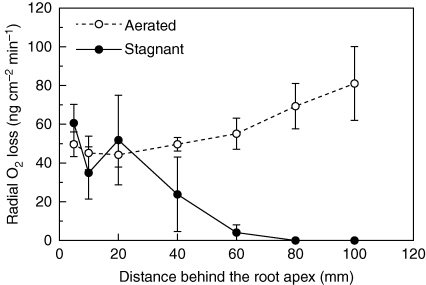
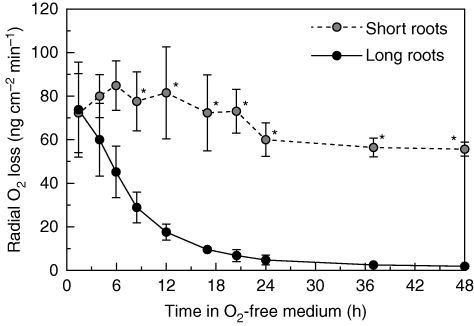
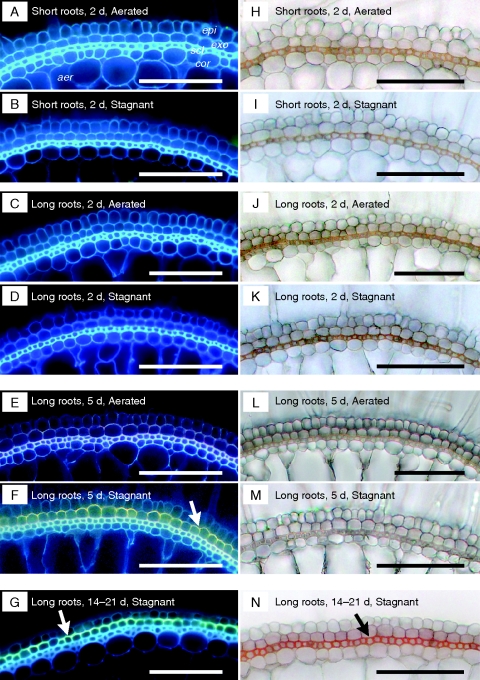
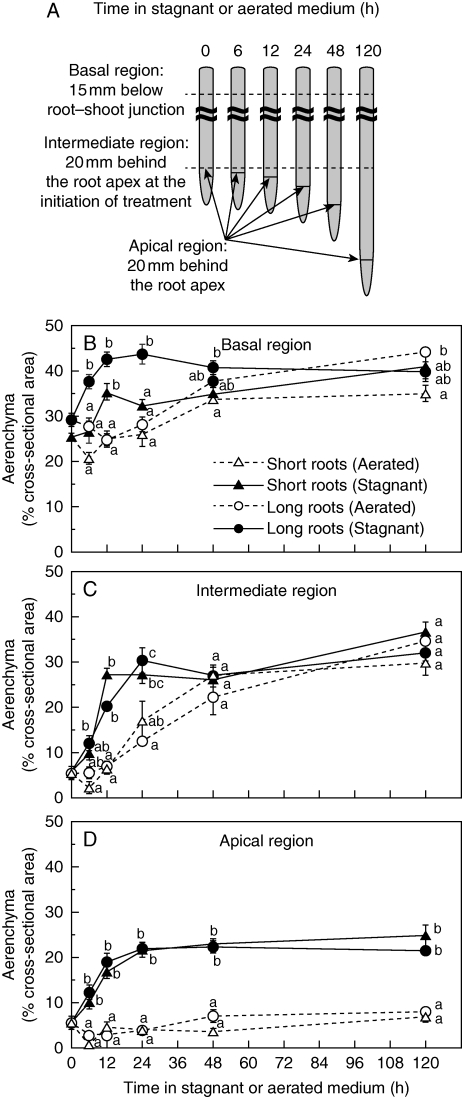
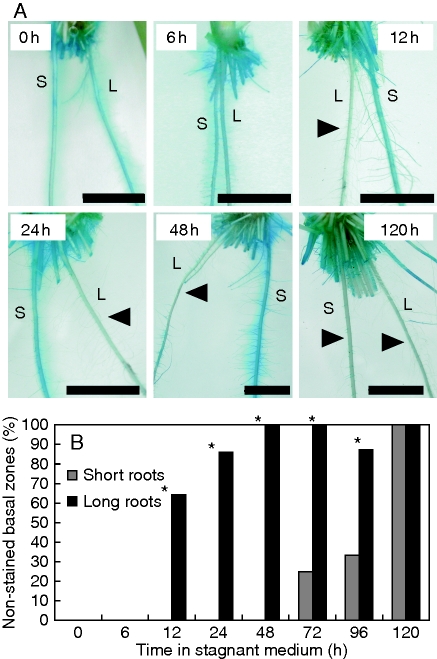
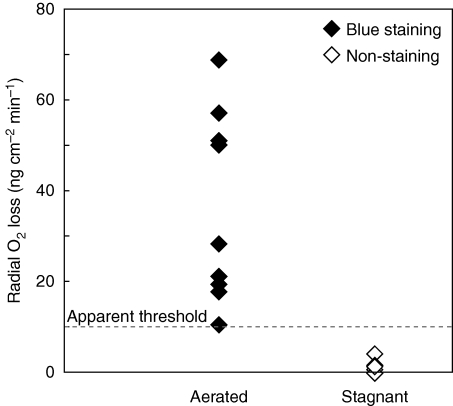
Key results: In stagnant medium, barrier to ROL formation commenced in long adventitious roots within a few hours and the barrier was well formed within 24 h. By contrast, barrier formation took longer than 48 h in short roots. The timing of enhancement of aerenchyma formation was the same in short and long roots. Comparison of ROL data and subsequent methylene blue staining determined the apparent ROL threshold for the dye method, and the dye method confirmed that barrier induction was faster for long roots than for short roots. Barrier formation might be related to deposition of new electron-dense materials in the cell walls at the peripheral side of the exodermis. Histochemical staining indicated suberin depositions were enhanced prior to increases in lignin.
Conclusions: As root length affected formation of the barrier to ROL, but not aerenchyma, these two acclimations are differentially regulated in roots of rice. Moreover, ROL barrier induction occurred before histochemically detectable changes in putative suberin and lignin deposits could be seen, whereas TEM showed deposition of new electron-dense materials in exodermal cell walls, so structural changes required for barrier functioning appear to be more subtle than previously described.
Figures
References
-
- Armstrong J, Armstrong W. Phragmites australis – A preliminary study of soil-oxidizing sites and internal gas transport pathways. New Phytologist. 1988;108:373–382.
-
- Armstrong J, Armstrong W. Rice and Phragmites: effects of organic acids on growth, root permeability, and radial oxygen loss to the rhizosphere. American Journal of Botany. 2001;88:1359–1370. - PubMed
-
- Armstrong J, Armstrong W, Beckett PM. Phragmites australis – Venturi- and humidity-induced pressure flows enhance rhizome aeration and rhizosphere oxidation. New Phytologist. 1992;120:197–207.
-
- Armstrong W. Radial oxygen losses from intact rice roots as affected by distance from the apex, respiration and waterlogging. Physiologia Plantarum. 1971;25:192–197.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
