

Binding of albumin promotes bacterial survival at the epithelial surface
- PMID: 21098039
- PMCID: PMC3024741
- DOI: 10.1074/jbc.M110.148171
Binding of albumin promotes bacterial survival at the epithelial surface
Abstract
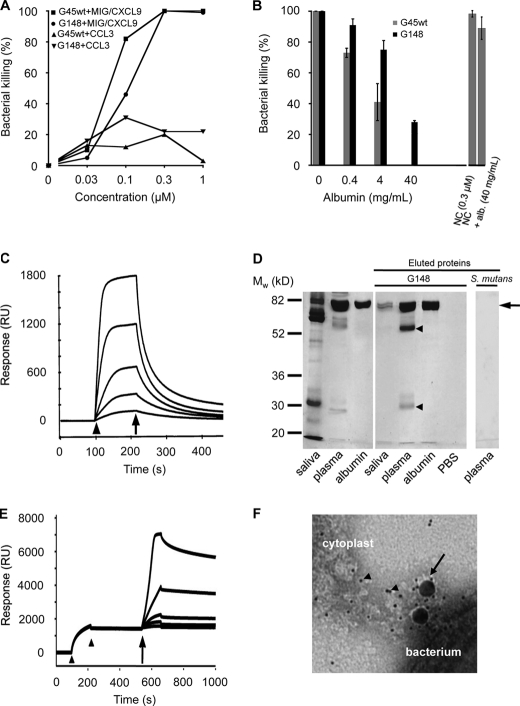
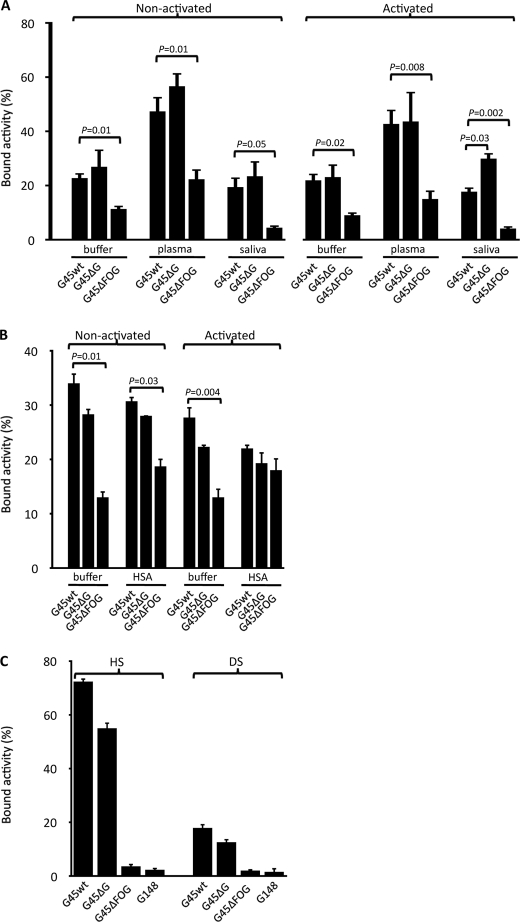
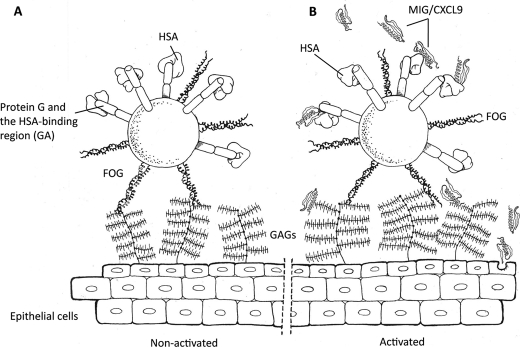
Human serum albumin (HSA) is the dominating protein in human plasma. Many bacterial species, especially streptococci, express surface proteins that bind HSA with high specificity and affinity, but the biological consequences of these protein-protein interactions are poorly understood. Group G streptococci (GGS), carrying the HSA-binding protein G, colonize the skin and the mucosa of the upper respiratory tract, mostly without causing disease. In the case of bacterial invasion, pro-inflammatory cytokines are released that activate the epithelium to produce antibacterial peptides, in particular the chemokine MIG/CXCL9. In addition, the inflammation causes capillary leakage and extravasation of HSA and other plasma proteins, environmental changes at the epithelial surface to which the bacteria need to respond. In this study, we found that GGS adsorbed HSA from both saliva and plasma via binding to protein G and that HSA bound to protein G bound and inactivated the antibacterial MIG/CXCL9 peptide. Another surface protein of GGS, FOG, was found to mediate adherence of the bacteria to pharyngeal epithelial cells through interaction with glycosaminoglycans. This adherence was not affected by activation of the epithelium with a combination of IFN-γ and TNF-α, leading to the production of MIG/CXCL9. However, at the activated epithelial surface, adherent GGS were protected against killing by MIG/CXCL9 through protein G-dependent HSA coating. The findings identify a previously unknown bacterial survival strategy that helps to explain the evolution of HSA-binding proteins among bacterial species of the normal human microbiota.
Figures

References
-
- Sylvetsky N., Raveh D., Schlesinger Y., Rudensky B., Yinnon A. M. (2002) Am. J. Med. 112, 622–626 - PubMed
-
- Sansonetti P. J. (2004) Nat. Rev. Immunol. 4, 953–964 - PubMed
-
- Hyland K. A., Brennan R., Olmsted S. B., Rojas E., Murphy E., Wang B., Cleary P. P. (2009) FEMS Immunol. Med. Microbiol. 55, 422–431 - PubMed
-
- Raeder R. H., Barker-Merrill L., Lester T., Boyle M. D., Metzger D. W. (2000) J. Infect. Dis. 181, 639–645 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
