

Transient activation of c-MYC expression is critical for efficient platelet generation from human induced pluripotent stem cells
- PMID: 21098095
- PMCID: PMC3005234
- DOI: 10.1084/jem.20100844
Transient activation of c-MYC expression is critical for efficient platelet generation from human induced pluripotent stem cells
Abstract
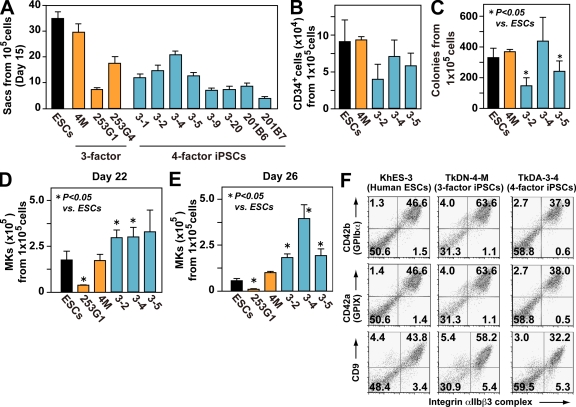
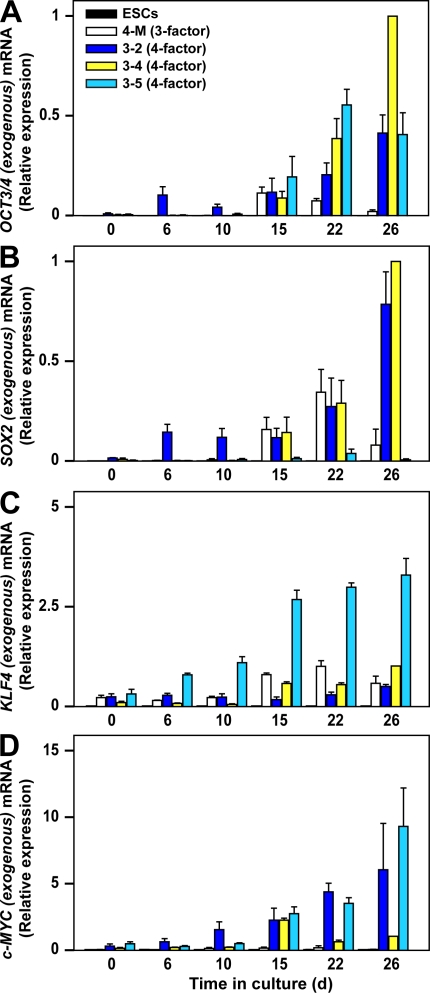
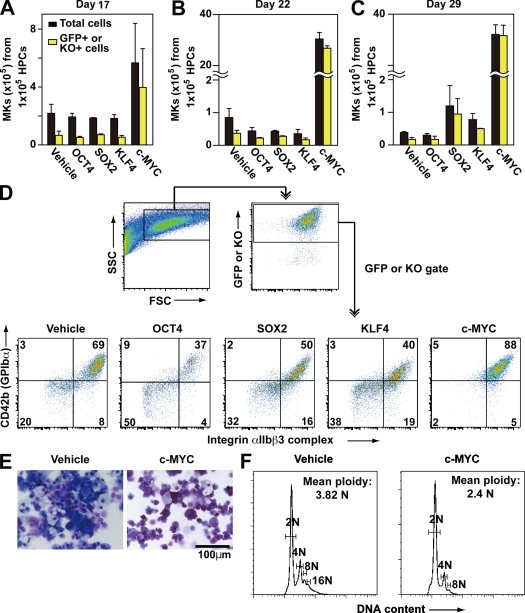
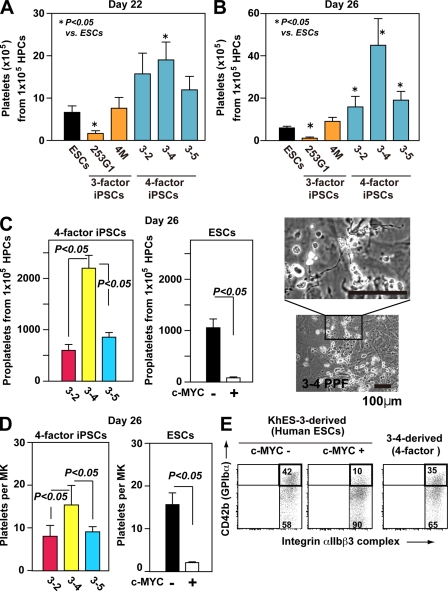
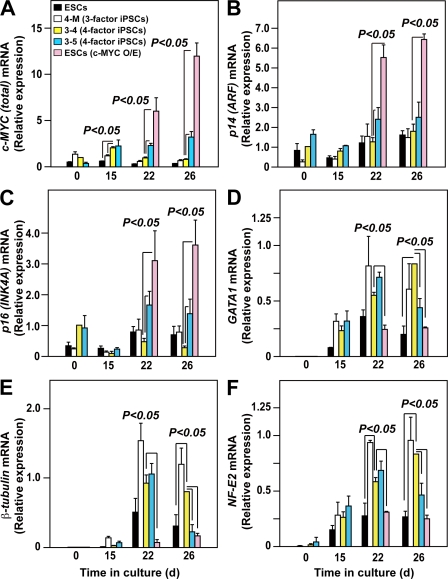
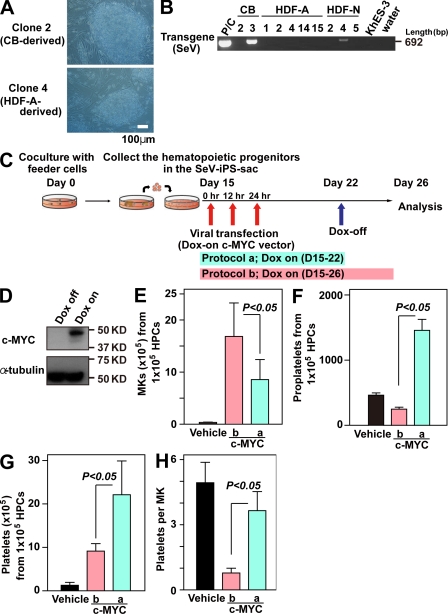
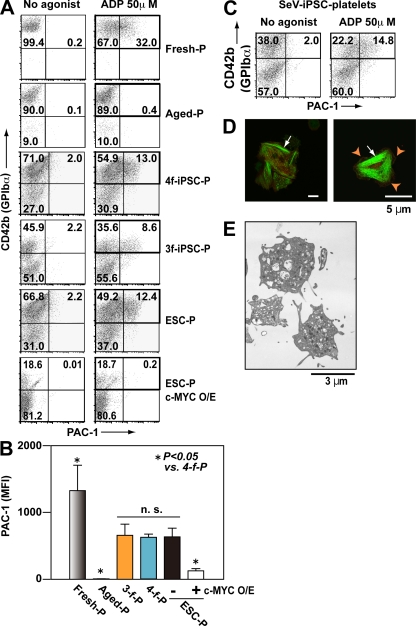
Human (h) induced pluripotent stem cells (iPSCs) are a potentially abundant source of blood cells, but how best to select iPSC clones suitable for this purpose from among the many clones that can be simultaneously established from an identical source is not clear. Using an in vitro culture system yielding a hematopoietic niche that concentrates hematopoietic progenitors, we show that the pattern of c-MYC reactivation after reprogramming influences platelet generation from hiPSCs. During differentiation, reduction of c-MYC expression after initial reactivation of c-MYC expression in selected hiPSC clones was associated with more efficient in vitro generation of CD41a(+)CD42b(+) platelets. This effect was recapitulated in virus integration-free hiPSCs using a doxycycline-controlled c-MYC expression vector. In vivo imaging revealed that these CD42b(+) platelets were present in thrombi after laser-induced vessel wall injury. In contrast, sustained and excessive c-MYC expression in megakaryocytes was accompanied by increased p14 (ARF) and p16 (INK4A) expression, decreased GATA1 expression, and impaired production of functional platelets. These findings suggest that the pattern of c-MYC expression, particularly its later decline, is key to producing functional platelets from selected iPSC clones.
Figures

Comment in
-
Induced pluripotent stem cell-derived human platelets: one step closer to the clinic.J Exp Med. 2010 Dec 20;207(13):2781-4. doi: 10.1084/jem.20102428. J Exp Med. 2010. PMID: 21173109 Free PMC article. Review.
References
-
- Askew D.S., Ashmun R.A., Simmons B.C., Cleveland J.L. 1991. Constitutive c-myc expression in an IL-3-dependent myeloid cell line suppresses cell cycle arrest and accelerates apoptosis. Oncogene. 6:1915–1922 - PubMed
-
- Chanprasert S., Geddis A.E., Barroga C., Fox N.E., Kaushansky K. 2006. Thrombopoietin (TPO) induces c-myc expression through a PI3K- and MAPK-dependent pathway that is not mediated by Akt, PKCzeta or mTOR in TPO-dependent cell lines and primary megakaryocytes. Cell. Signal. 18:1212–1218 10.1016/j.cellsig.2005.09.010 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
