

An invasive podosome-like structure promotes fusion pore formation during myoblast fusion
- PMID: 21098115
- PMCID: PMC2995175
- DOI: 10.1083/jcb.201006006
An invasive podosome-like structure promotes fusion pore formation during myoblast fusion
Abstract
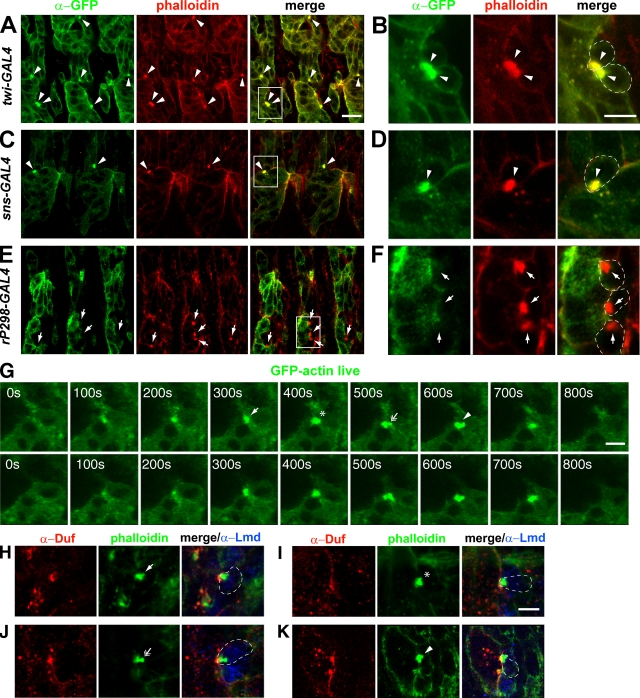
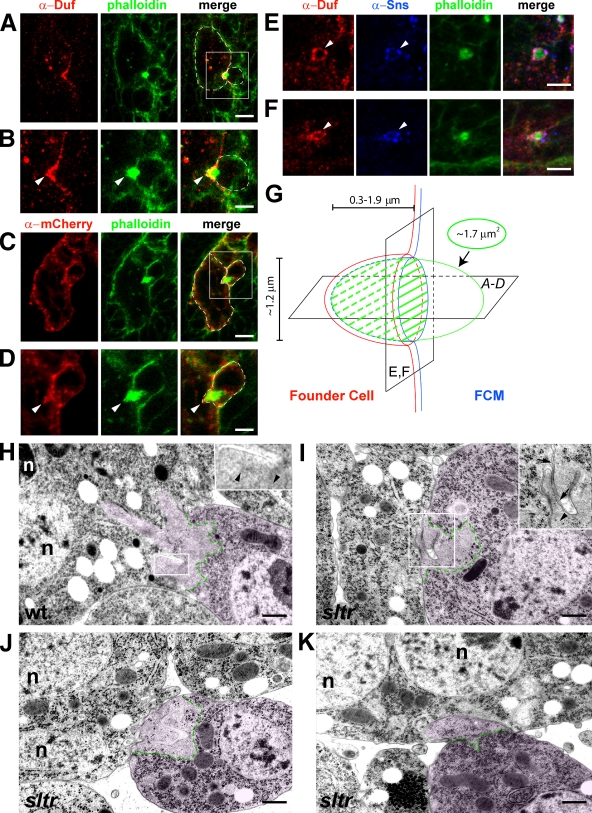
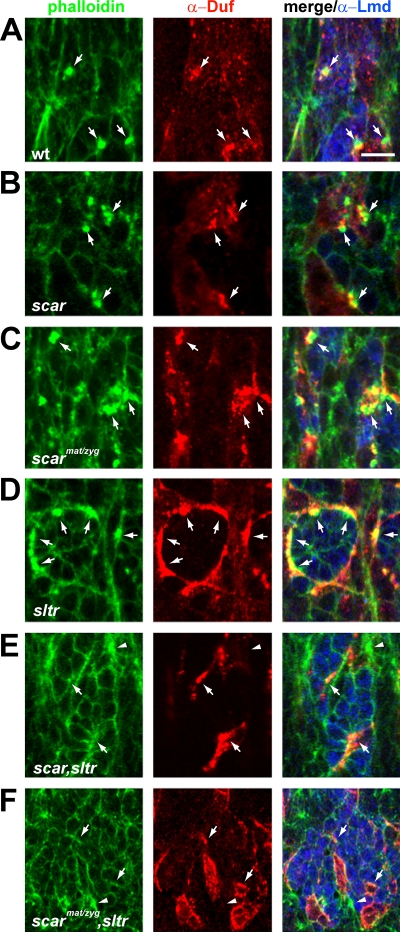
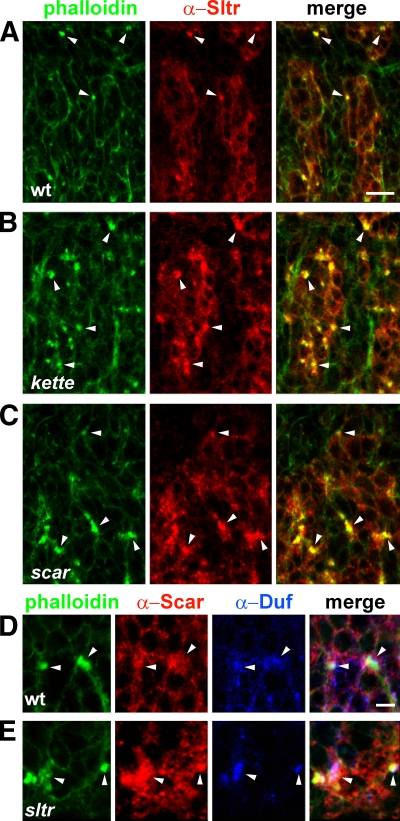
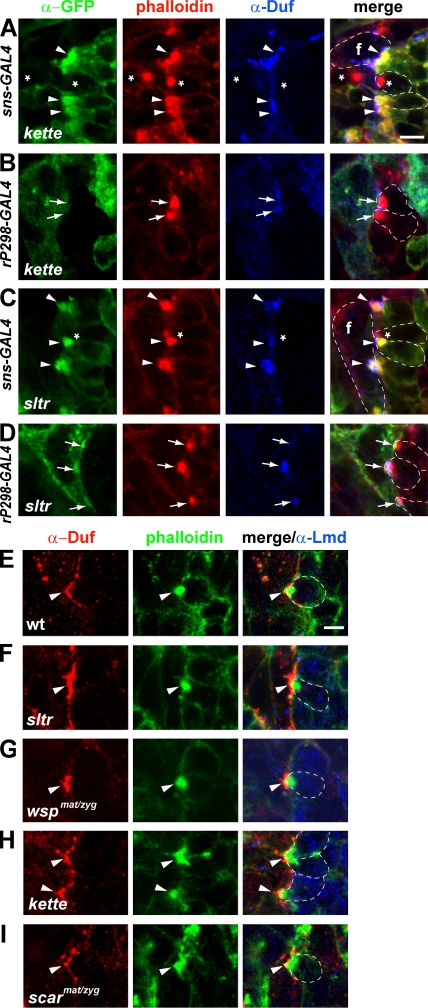
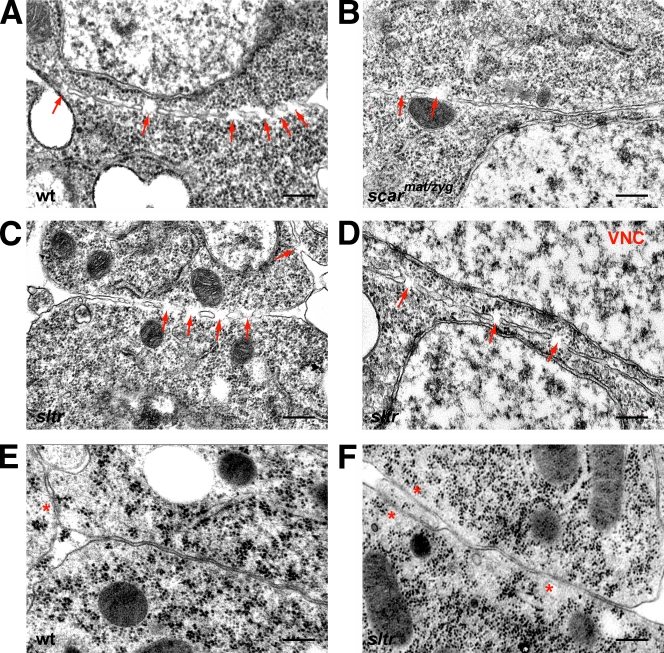
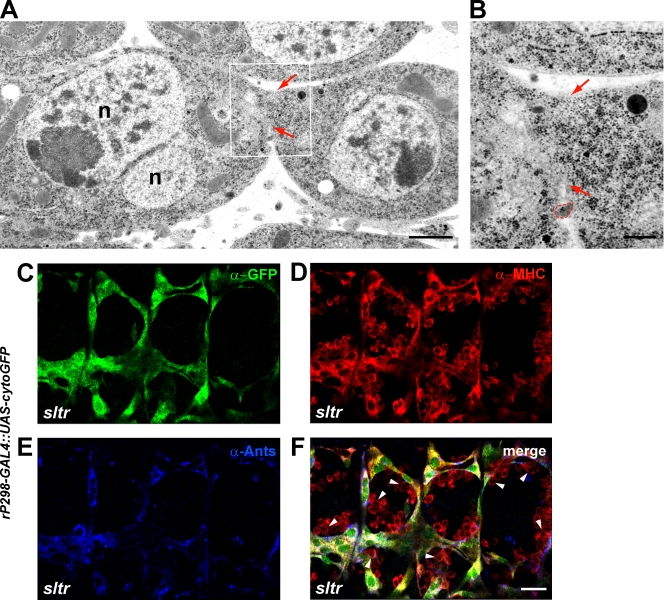
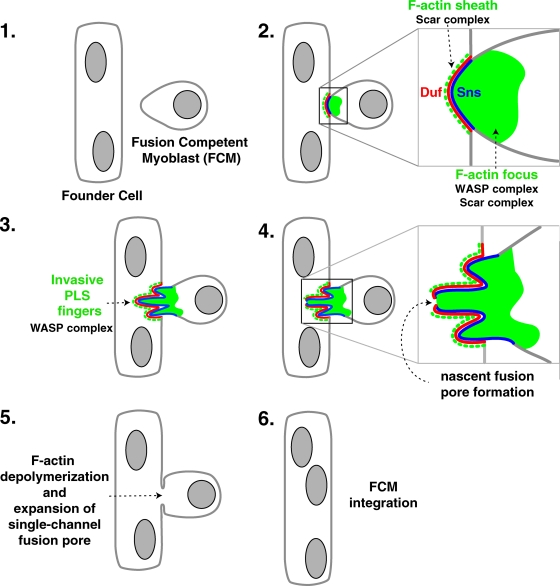
Recent studies in Drosophila have implicated actin cytoskeletal remodeling in myoblast fusion, but the cellular mechanisms underlying this process remain poorly understood. Here we show that actin polymerization occurs in an asymmetric and cell type-specific manner between a muscle founder cell and a fusion-competent myoblast (FCM). In the FCM, a dense F-actin-enriched focus forms at the site of fusion, whereas a thin sheath of F-actin is induced along the apposing founder cell membrane. The FCM-specific actin focus invades the apposing founder cell with multiple finger-like protrusions, leading to the formation of a single-channel macro fusion pore between the two muscle cells. Two actin nucleation-promoting factors of the Arp2/3 complex, WASP and Scar, are required for the formation of the F-actin foci, whereas WASP but not Scar promotes efficient foci invasion. Our studies uncover a novel invasive podosome-like structure (PLS) in a developing tissue and reveal a previously unrecognized function of PLSs in facilitating cell membrane juxtaposition and fusion.
Figures
References
-
- Artero R.D., Castanon I., Baylies M.K. 2001. The immunoglobulin-like protein Hibris functions as a dose-dependent regulator of myoblast fusion and is differentially controlled by Ras and Notch signaling. Development. 128:4251–4264 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
