

DEAH-box ATPase Prp16 has dual roles in remodeling of the spliceosome in catalytic steps
- PMID: 21098140
- PMCID: PMC3004056
- DOI: 10.1261/rna.2459611
DEAH-box ATPase Prp16 has dual roles in remodeling of the spliceosome in catalytic steps
Abstract
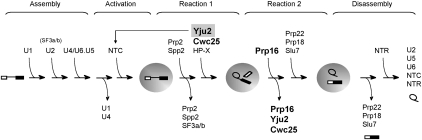
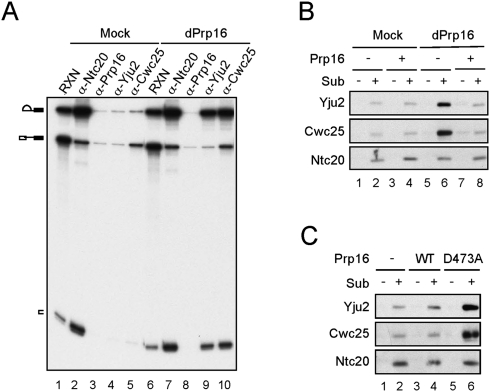
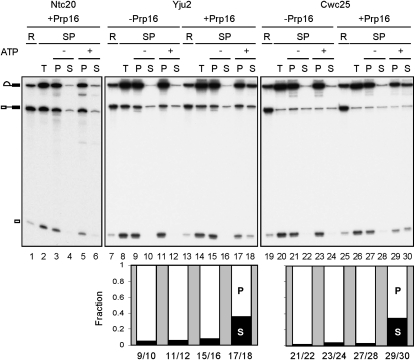
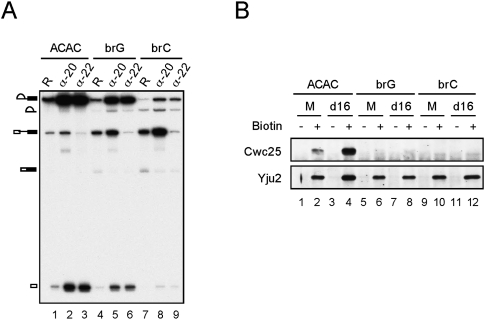
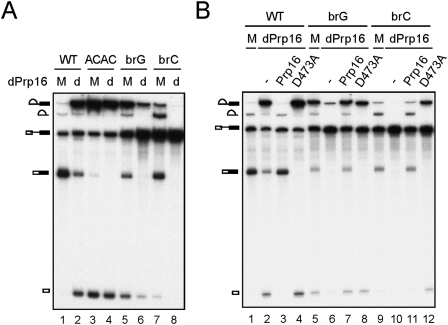
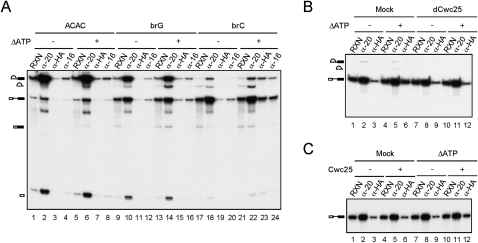
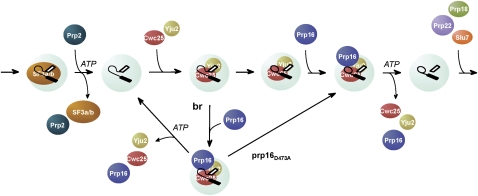
The assembly of the spliceosome involves dynamic rearrangements of interactions between snRNAs, protein components, and the pre-mRNA substrate. DExD/H-box ATPases are required to mediate structural changes of the spliceosome, utilizing the energy of ATP hydrolysis. Two DExD/H-box ATPases are required for the catalytic steps of the splicing pathway, Prp2 for the first step and Prp16 for the second step, both belonging to the DEAH subgroup of the protein family. The detailed mechanism of their action was not well understood until recently, when Prp2 was shown to be required for the release of U2 components SF3a and SF3b, presumably to allow the binding of Cwc25 to promote the first transesterification reaction. We show here that Cwc25 and Yju2 are released after the reaction in Prp16- and ATP-dependent manners, possibly to allow for the binding of Prp22, Prp18, and Slu7 to promote the second catalytic reaction. The binding of Cwc25 to the spliceosome is destabilized by mutations at the branchpoint sequence, suggesting that Cwc25 may bind to the branch site. We also show that Prp16 has an ATP-independent role in the first catalytic step, in addition to its known role in the second step. In the absence of ATP, Prp16 stabilizes the binding of Cwc25 to the spliceosome formed with branchpoint mutated pre-mRNAs to facilitate their splicing. Our results uncovered novel functions of Prp16 in both catalytic steps, and provide mechanistic insights into splicing catalysis.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases