

The role of alpha-glucosidase in germinating barley grains
- PMID: 21098673
- PMCID: PMC3032477
- DOI: 10.1104/pp.110.168328
The role of alpha-glucosidase in germinating barley grains
Abstract
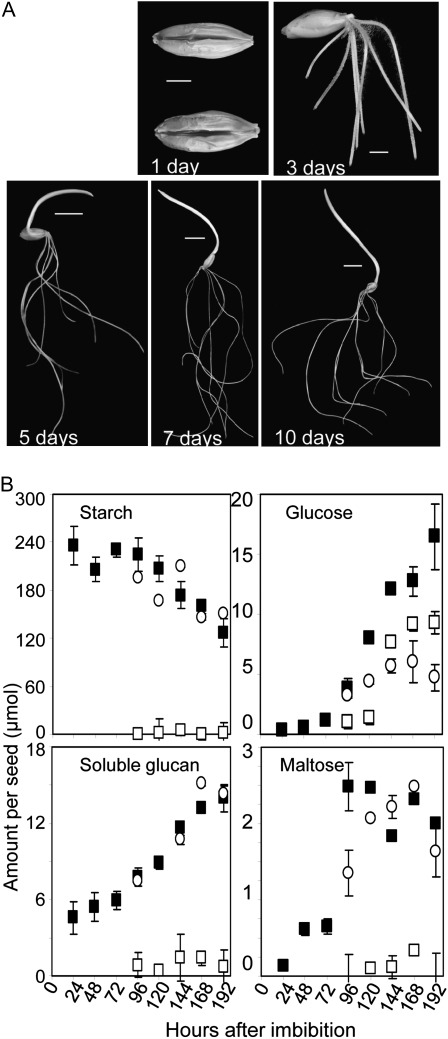
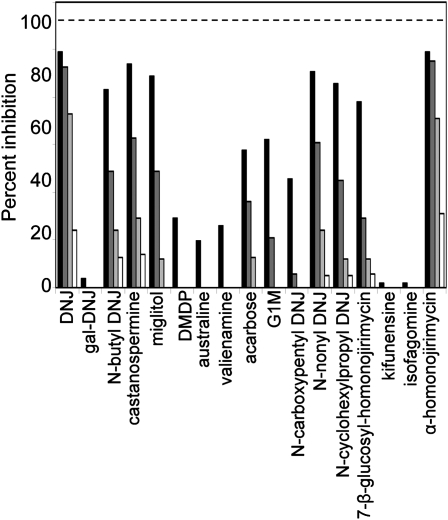
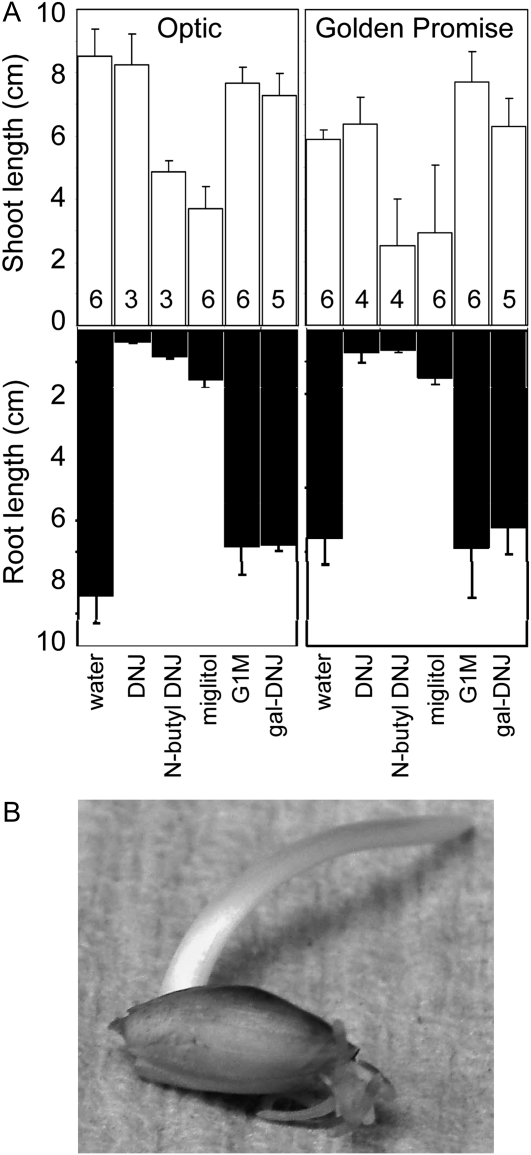
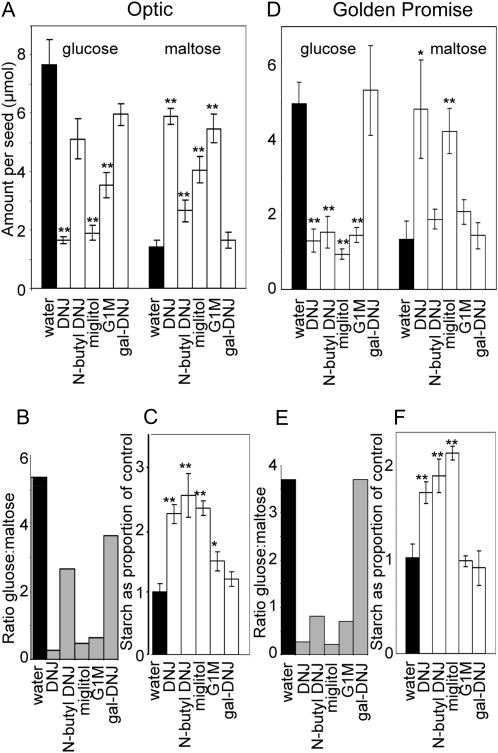
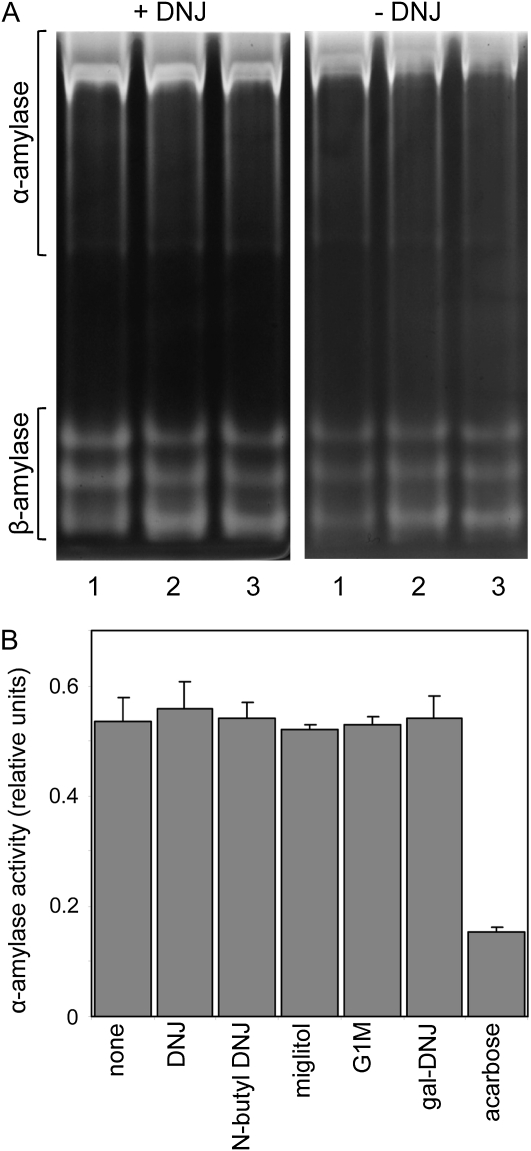
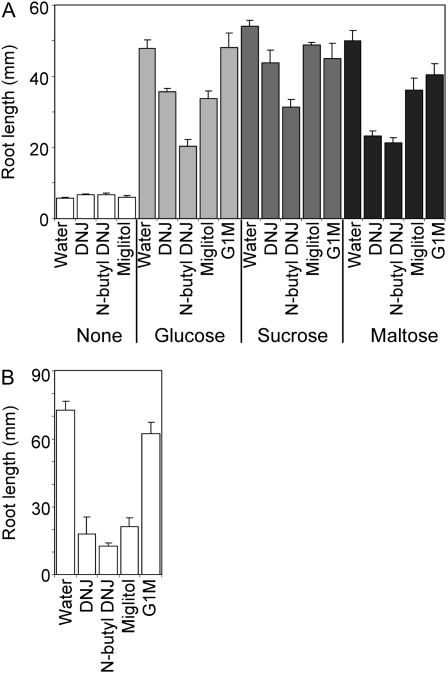
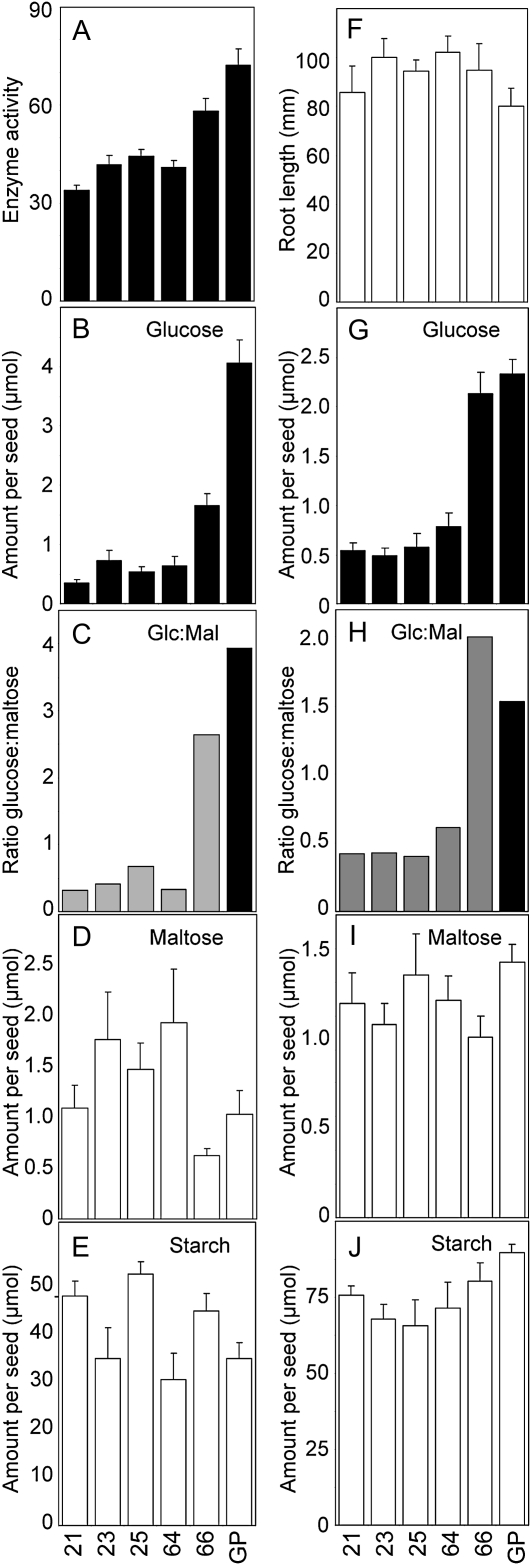
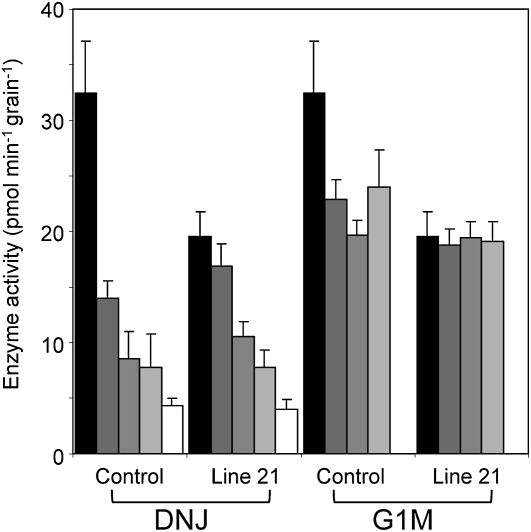
The importance of α-glucosidase in the endosperm starch metabolism of barley (Hordeum vulgare) seedlings is poorly understood. The enzyme converts maltose to glucose (Glc), but in vitro studies indicate that it can also attack starch granules. To discover its role in vivo, we took complementary chemical-genetic and reverse-genetic approaches. We identified iminosugar inhibitors of a recombinant form of an α-glucosidase previously discovered in barley endosperm (ALPHA-GLUCOSIDASE97 [HvAGL97]), and applied four of them to germinating grains. All four decreased the Glc-to-maltose ratio in the endosperm 10 d after imbibition, implying inhibition of maltase activity. Three of the four inhibitors also reduced starch degradation and seedling growth, but the fourth did not affect these parameters. Inhibition of starch degradation was apparently not due to inhibition of amylases. Inhibition of seedling growth was primarily a direct effect of the inhibitors on roots and coleoptiles rather than an indirect effect of the inhibition of endosperm metabolism. It may reflect inhibition of glycoprotein-processing glucosidases in these organs. In transgenic seedlings carrying an RNA interference silencing cassette for HvAgl97, α-glucosidase activity was reduced by up to 50%. There was a large decrease in the Glc-to-maltose ratio in these lines but no effect on starch degradation or seedling growth. Our results suggest that the α-glucosidase HvAGL97 is the major endosperm enzyme catalyzing the conversion of maltose to Glc but is not required for starch degradation. However, the effects of three glucosidase inhibitors on starch degradation in the endosperm indicate the existence of unidentified glucosidase(s) required for this process.
Figures
References
-
- Al Kazaz M, Desseaux V, Marchis-Mouren G, Prodanov E, Santimone M. (1998) The mechanism of porcine pancreatic α-amylase: inhibition of maltopentaose hydrolysis by acarbose, maltose and maltotriose. Eur J Biochem 252: 100–107 - PubMed
-
- Arai M, Sumida M, Fukuhara K, Kainosho M, Murao S. (1986) Isolation and characterization of amylase inhibitors, (glucose)n·deoxynojirimycin. Agric Biol Chem 50: 639–644
-
- Asano N, Kato A, Kizu H, Matsui K, Watson AA, Nash RJ. (1996) Calystegine B4, a novel trehalase inhibitor from Scopolia japonica. Carbohydr Res 293: 195–204 - PubMed
-
- Bamforth CW. (2009) Current perspectives on the role of enzymes in brewing. J Cereal Sci 50: 353–357
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
