

αv Integrin expression by DCs is required for Th17 cell differentiation and development of experimental autoimmune encephalomyelitis in mice
- PMID: 21099114
- PMCID: PMC2993596
- DOI: 10.1172/JCI43796
αv Integrin expression by DCs is required for Th17 cell differentiation and development of experimental autoimmune encephalomyelitis in mice
Abstract
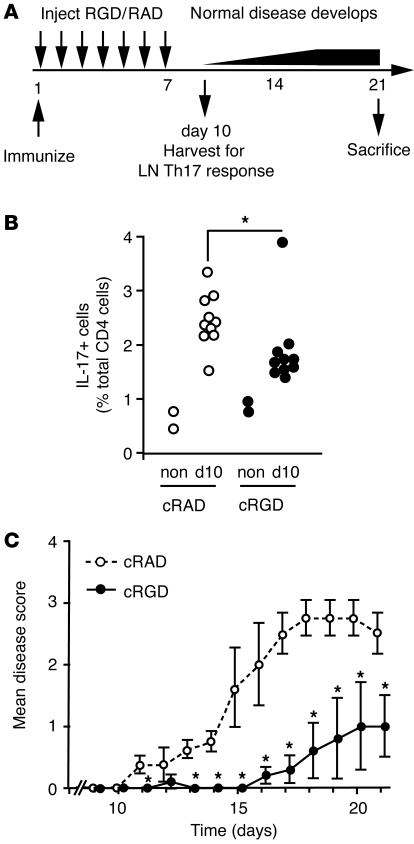
Th17 cells are a distinct lineage of T helper cells that protect the body from bacterial and fungal infection. However, Th17 cells also contribute to inflammatory and autoimmune disorders such as multiple sclerosis. Th17 cell generation requires exposure of naive T cells to the cytokine TGF-β in combination with proinflammatory cytokines. Here we show that differentiation of Th17 cells is also critically dependent on αv integrins. In mice, lack of integrin αv in the immune system resulted in loss of Th17 cells in the intestine and lymphoid tissues. It also led to protection from experimental autoimmune encephalomyelitis (EAE). Further analysis indicated that αv integrins on DCs activated latent TGF-β during T cell stimulation and thereby promoted differentiation of Th17 cells. Furthermore, pharmacologic inhibition of αv integrins using cyclic RGD peptides blocked TGF-β activation and Th17 cell generation in vitro and protected mice from EAE. These data demonstrate that activation of TGF-β by αv-expressing myeloid cells may be a critical step in the generation of Th17 cells and suggest that αv integrins could be therapeutic targets in autoimmune disease.
Figures
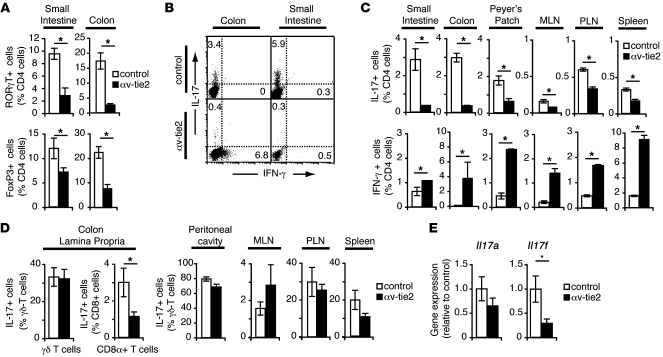
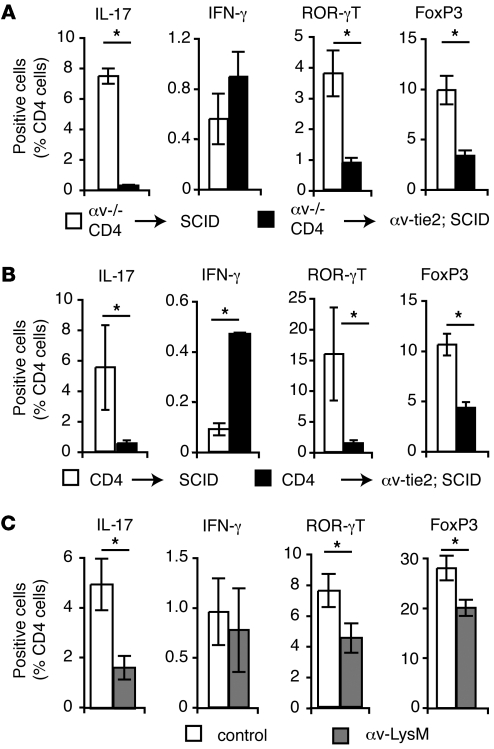
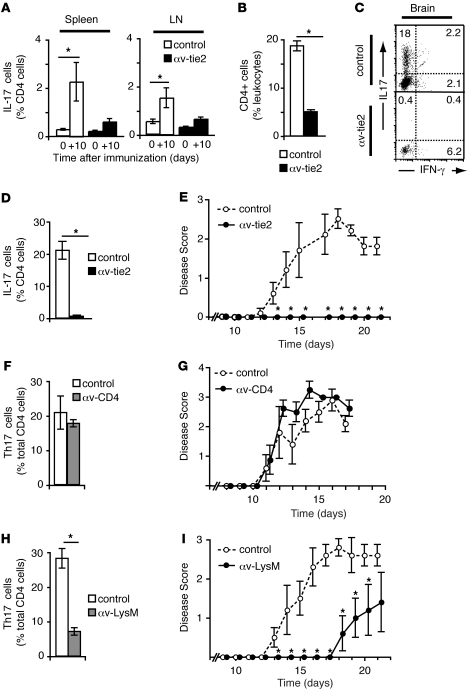
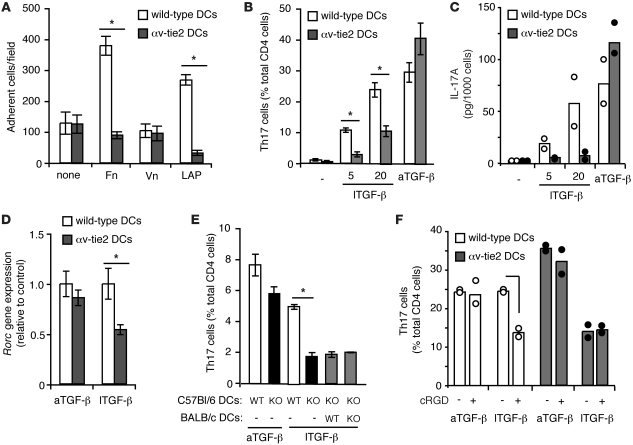
Comment in
-
Integral role of integrins in Th17 development.J Clin Invest. 2010 Dec;120(12):4185-7. doi: 10.1172/JCI45450. Epub 2010 Nov 22. J Clin Invest. 2010. PMID: 21099101 Free PMC article.
References
-
- Steinman L. A brief history of T(H)17, the first major revision in the T(H)1/T(H)2 hypothesis of T cell-mediated tissue damage. Nat Med. 2007;13(2):139–145. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
