

Phosphorylation of the ryanodine receptor mediates the cardiac fight or flight response in mice
- PMID: 21099118
- PMCID: PMC2993575
- DOI: 10.1172/JCI32726
Phosphorylation of the ryanodine receptor mediates the cardiac fight or flight response in mice
Abstract
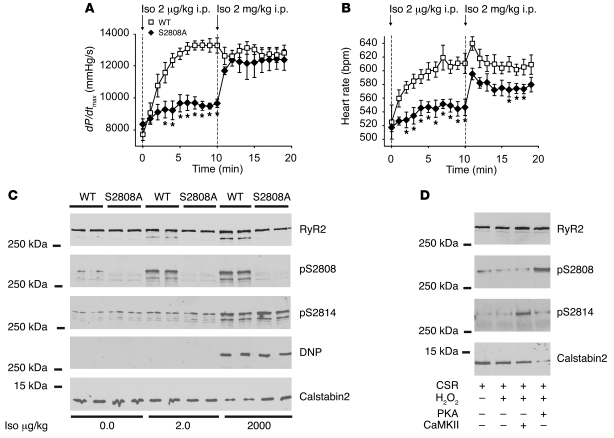
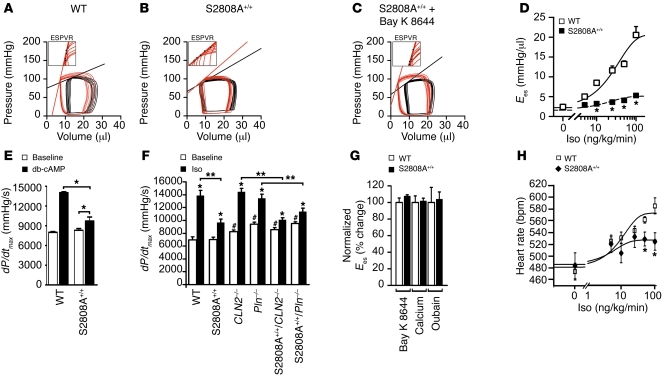
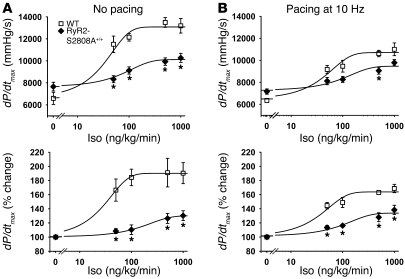
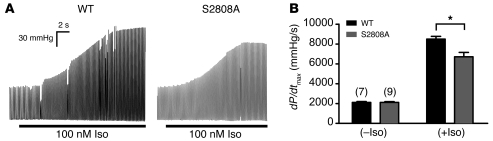
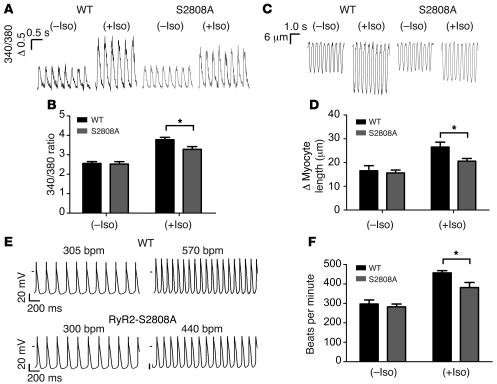
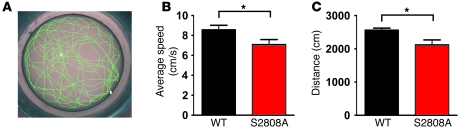
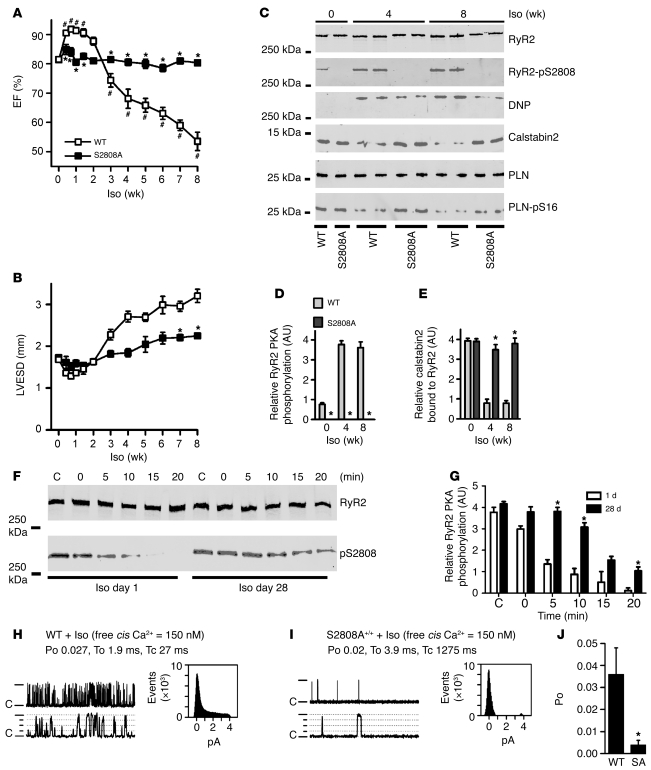
During the classic "fight-or-flight" stress response, sympathetic nervous system activation leads to catecholamine release, which increases heart rate and contractility, resulting in enhanced cardiac output. Catecholamines bind to β-adrenergic receptors, causing cAMP generation and activation of PKA, which phosphorylates multiple targets in cardiac muscle, including the cardiac ryanodine receptor/calcium release channel (RyR2) required for muscle contraction. PKA phosphorylation of RyR2 enhances channel activity by sensitizing the channel to cytosolic calcium (Ca²+). Here, we found that mice harboring RyR2 channels that cannot be PKA phosphorylated (referred to herein as RyR2-S2808A+/+ mice) exhibited blunted heart rate and cardiac contractile responses to catecholamines (isoproterenol). The isoproterenol-induced enhancement of ventricular myocyte Ca²+ transients and fractional shortening (contraction) and the spontaneous beating rate of sinoatrial nodal cells were all blunted in RyR2-S2808A+/+ mice. The blunted cardiac response to catecholamines in RyR2-S2808A+/+ mice resulted in impaired exercise capacity. RyR2-S2808A+/+ mice were protected against chronic catecholaminergic-induced cardiac dysfunction. These studies identify what we believe to be new roles for PKA phosphorylation of RyR2 in both the heart rate and contractile responses to acute catecholaminergic stimulation.
Figures
Comment in
-
Is ryanodine receptor phosphorylation key to the fight or flight response and heart failure?J Clin Invest. 2010 Dec;120(12):4197-203. doi: 10.1172/JCI45251. Epub 2010 Nov 22. J Clin Invest. 2010. PMID: 21099119 Free PMC article.
References
-
- Vatner SF. Sympathetic mechanisms regulating myocardial contractility in concious animals. In: Fozzard HA, Haber E, eds.The Heart and Cardiovascular System . New York, New York, USA: Raven Press, Ltd; 1992:1709–1728.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
