

Activation of pulmonary dendritic cells and Th2-type inflammatory responses on instillation of engineered, environmental diesel emission source or ambient air pollutant particles in vivo
- PMID: 21099199
- PMCID: PMC3072202
- DOI: 10.1159/000321725
Activation of pulmonary dendritic cells and Th2-type inflammatory responses on instillation of engineered, environmental diesel emission source or ambient air pollutant particles in vivo
Abstract
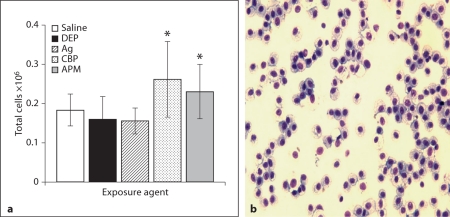
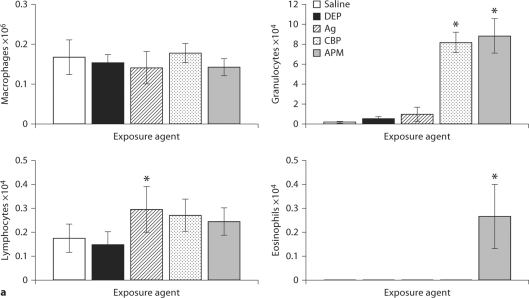
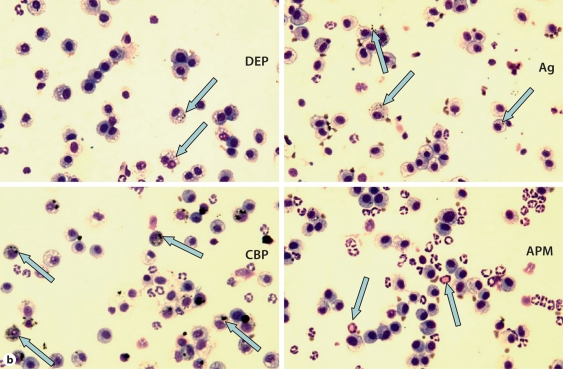
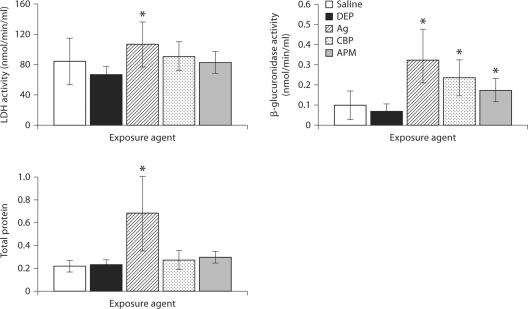
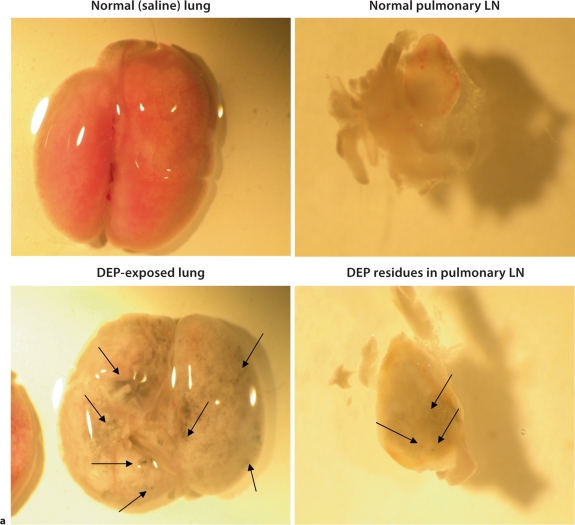
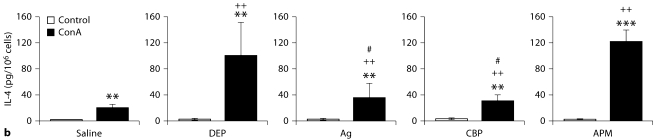
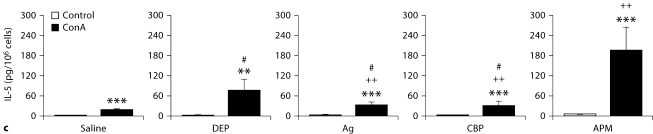
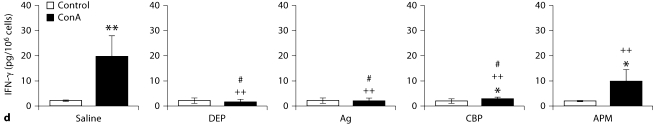
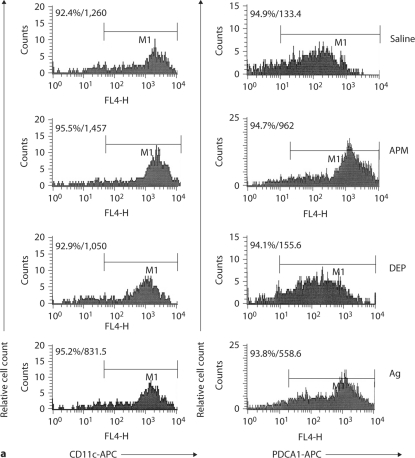
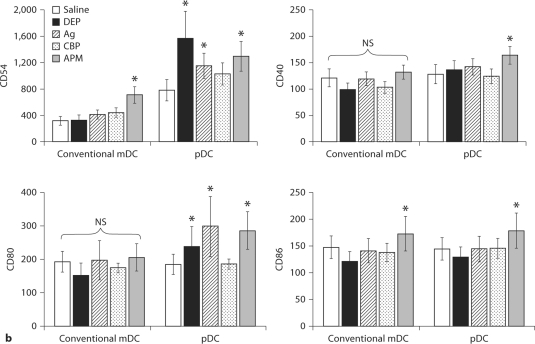
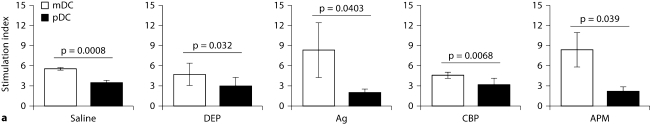
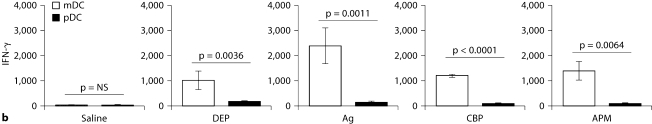
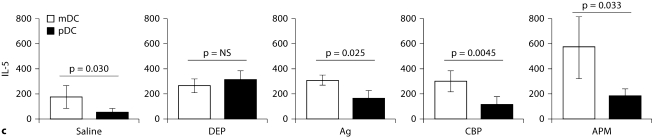
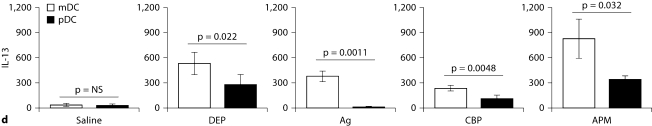
The biological effects of acute particulate air pollution exposure in host innate immunity remain obscure and have relied largely on in vitro models. We hypothesized that single acute exposure to ambient or engineered particulate matter (PM) in the absence of other secondary stimuli would activate lung dendritic cells (DC) in vivo and provide information on the early immunological events of PM exposure and DC activation in a mouse model naïve to prior PM exposure. Activation of purified lung DC was studied following oropharyngeal instillation of ambient particulate matter (APM). We compared the effects of APM exposure with that of diesel-enriched PM (DEP), carbon black particles (CBP) and silver nanoparticles (AgP). We found that PM species induced variable cellular infiltration in the lungs and only APM exposure induced eosinophilic infiltration. Both APM and DEP activated pulmonary DC and promoted a Th2-type cytokine response from naïve CD4+ T cells ex vivo. Cultures of primary peribronchial lymph node cells from mice exposed to APM and DEP also displayed a Th2-type immune response ex vivo. We conclude that exposure of the lower airway to various PM species induces differential immunological responses and immunomodulation of DC subsets. Environmental APM and DEP activated DC in vivo and provoked a Th2 response ex vivo. By contrast, CBP and AgP induced altered lung tissue barrier integrity but failed to stimulate CD4+ T cells as effectively. Our work suggests that respirable pollutants activate the innate immune response with enhanced DC activation, pulmonary inflammation and Th2-immune responsiveness.
Copyright © 2010 S. Karger AG, Basel.
Figures

Similar articles
-
Diesel-enriched particulate matter functionally activates human dendritic cells.Am J Respir Cell Mol Biol. 2007 Dec;37(6):706-19. doi: 10.1165/rcmb.2007-0199OC. Epub 2007 Jul 13. Am J Respir Cell Mol Biol. 2007. PMID: 17630318 Free PMC article.
-
Effects of diesel exhaust particles on antigen-presenting cells and antigen-specific Th immunity in mice.Exp Biol Med (Maywood). 2009 Feb;234(2):200-9. doi: 10.3181/0809-RM-285. Epub 2008 Dec 8. Exp Biol Med (Maywood). 2009. PMID: 19064938
-
Ambient particulate matter directs nonclassic dendritic cell activation and a mixed TH1/TH2-like cytokine response by naive CD4+ T cells.J Allergy Clin Immunol. 2007 Feb;119(2):488-97. doi: 10.1016/j.jaci.2006.10.022. Epub 2006 Dec 21. J Allergy Clin Immunol. 2007. PMID: 17187851
-
The dual effect of the particulate and organic components of diesel exhaust particles on the alteration of pulmonary immune/inflammatory responses and metabolic enzymes.J Environ Sci Health C Environ Carcinog Ecotoxicol Rev. 2002 Nov;20(2):117-47. doi: 10.1081/GNC-120016202. J Environ Sci Health C Environ Carcinog Ecotoxicol Rev. 2002. PMID: 12515672 Review.
-
Effect of diesel exhaust particulate (DEP) on immune responses: contributions of particulate versus organic soluble components.J Toxicol Environ Health A. 2004 Feb 13;67(3):221-31. doi: 10.1080/15287390490266891. J Toxicol Environ Health A. 2004. PMID: 14681077 Review.
Cited by
-
Titanium dioxide nanoparticles exaggerate respiratory syncytial virus-induced airway epithelial barrier dysfunction.Am J Physiol Lung Cell Mol Physiol. 2020 Sep 1;319(3):L481-L496. doi: 10.1152/ajplung.00104.2020. Epub 2020 Jul 8. Am J Physiol Lung Cell Mol Physiol. 2020. PMID: 32640839 Free PMC article.
-
Preventing carbon nanoparticle-induced lung inflammation reduces antigen-specific sensitization and subsequent allergic reactions in a mouse model.Part Fibre Toxicol. 2015 Jul 4;12:20. doi: 10.1186/s12989-015-0093-5. Part Fibre Toxicol. 2015. PMID: 26141115 Free PMC article.
-
Psychological stress: a social pollutant that may enhance environmental risk.Am J Respir Crit Care Med. 2011 Oct 1;184(7):752-4. doi: 10.1164/rccm.201106-1139ED. Am J Respir Crit Care Med. 2011. PMID: 21965012 Free PMC article. No abstract available.
-
A work group report on ultrafine particles (American Academy of Allergy, Asthma & Immunology): Why ambient ultrafine and engineered nanoparticles should receive special attention for possible adverse health outcomes in human subjects.J Allergy Clin Immunol. 2016 Aug;138(2):386-96. doi: 10.1016/j.jaci.2016.02.023. Epub 2016 Apr 6. J Allergy Clin Immunol. 2016. PMID: 27130856 Free PMC article. Review.
-
Mechanisms linking traffic-related air pollution and atherosclerosis.Curr Opin Pulm Med. 2012 Mar;18(2):155-60. doi: 10.1097/MCP.0b013e32834f210a. Curr Opin Pulm Med. 2012. PMID: 22189455 Free PMC article. Review.
References
-
- Cohn L, Elias JA, Chupp GL. Asthma: mechanisms of disease persistence and progression. Annu Rev Immunol. 2004;22:789–815. - PubMed
-
- Nel AE, Diaz-Sanchez D, Ng D, Hiura T, Saxon A. Enhancement of allergic inflammation by the interaction between diesel exhaust particles and the immune system. J Allergy Clin Immunol. 2008;102:539–554. - PubMed
-
- Wills-Karp M. Immunologic basis of antigen-induced airway hyperresponsiveness. Annu Rev Immunol. 1999;17:255–281. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
