

Characterization of the gating brake in the I-II loop of CaV3 T-type calcium channels
- PMID: 21099341
- PMCID: PMC3052245
- DOI: 10.4161/chan.4.6.12889
Characterization of the gating brake in the I-II loop of CaV3 T-type calcium channels
Abstract
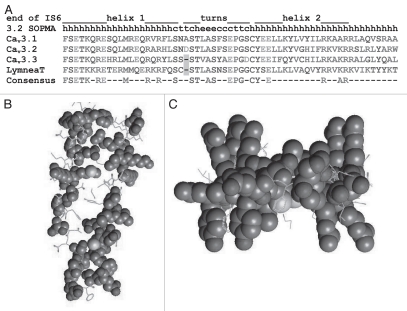
Our interest was drawn to the I-II loop of Cav3 channels for two reasons: one, transfer of the I-II loop from a high voltage-activated channel (Cav2.2) to a low voltage-activated channel (Cav3.1) unexpectedly produced an ultra-low voltage activated channel; and two, sequence variants of the I-II loop found in childhood absence epilepsy patients altered channel gating and increased surface expression of Cav3.2 channels. To determine the roles of this loop we have studied the structure of the loop and the biophysical consequences of altering its structure. Deletions localized the gating brake to the first 62 amino acids after IS6 in all three Cav3 channels, establishing the evolutionary conservation of this region and its function. Circular dichroism was performed on a purified fragment of the I-II loop from Cav3.2 to reveal a high α-helical content. De novo computer modeling predicted the gating brake formed a helix-loop-helix structure. This model was tested by replacing the helical regions with poly-proline-glycine (PGPGPG), which introduces kinks and flexibility. These mutations had profound effects on channel gating, shifting both steady-state activation and inactivation curves, as well as accelerating channel kinetics. Mutations designed to preserve the helical structure (poly-alanine, which forms α-helices) had more modest effects. Taken together, we conclude the second helix of the gating brake establishes important contacts with the gating machinery, thereby stabilizing a closed state of T-channels, and that this interaction is disrupted by depolarization, allowing the S6 segments to spread open and Ca (2+) ions to flow through.
Figures
References
-
- Zamponi GW, Bourinet E, Nelson D, Nargeot J, Snutch TP. Crosstalk between G proteins and protein kinase C mediated by the calcium channel a1 subunit. Nature. 1997;385:442–446. - PubMed
-
- Arias JM, Murbartián J, Vitko I, Lee JH, Perez-Reyes E. Transfer of β subunit regulation from high to low voltage-gated Ca2+ channels. FEBS Lett. 2005;579:3907–3912. - PubMed
-
- Raybaud A, Dodier Y, Bissonnette P, Simoes M, Bichet DG, Sauve R, et al. The role of the GX9GX3G motif in the gating of high voltage-activated Ca2+ channels. J Biol Chem. 2006;281:39424–39436. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources