

Review
doi: 10.1038/nrm3010.
Asymmetric cell division: recent developments and their implications for tumour biology
Affiliations
- PMID: 21102610
- PMCID: PMC3941022
- DOI: 10.1038/nrm3010
Item in Clipboard
Review
Asymmetric cell division: recent developments and their implications for tumour biology
Nat Rev Mol Cell Biol.
2010 Dec.
Abstract
The ability of cells to divide asymmetrically is essential for generating diverse cell types during development. The past 10 years have seen tremendous progress in our understanding of this important biological process. We have learned that localized phosphorylation events are responsible for the asymmetric segregation of cell fate determinants in mitosis and that centrosomes and microtubules play important parts in this process. The relevance of asymmetric cell division for stem cell biology has added a new dimension to the field, and exciting connections between asymmetric cell division and tumorigenesis have begun to emerge.
Figures
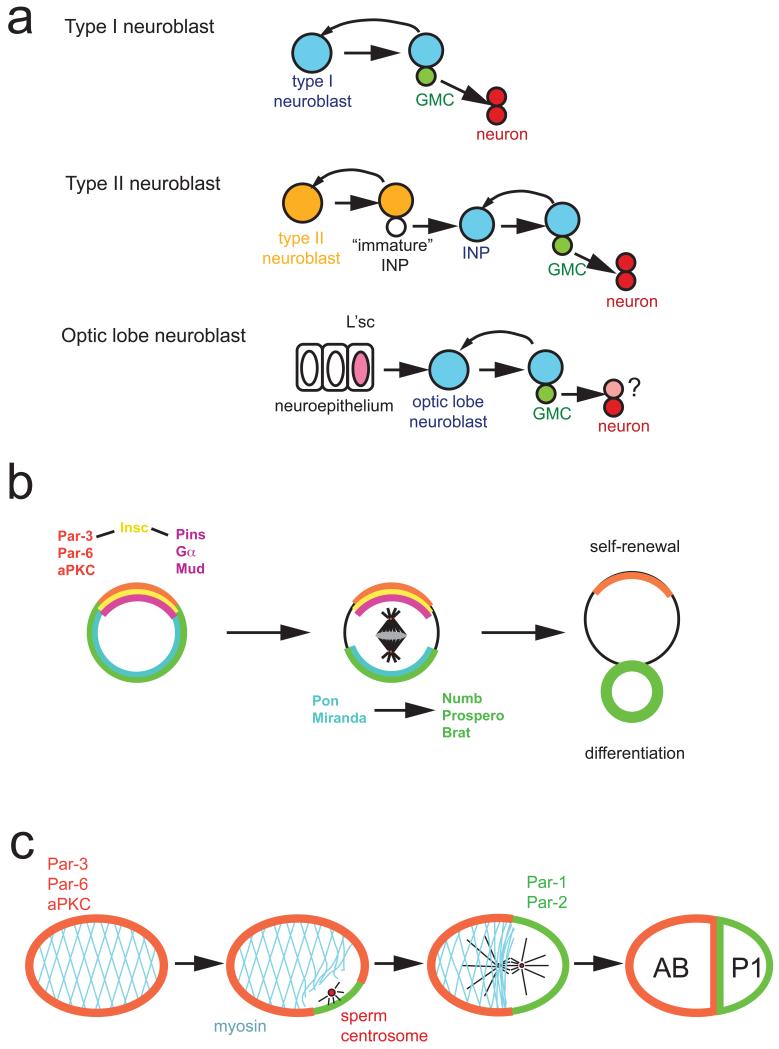
a ∣ Drosophila type 1 neuroblasts (top, blue) divide asymmetrically into one neuroblast and a ganglion mother cell (GMC, green). The neuroblast self-renews while the GMC divides terminally into two neurons. Type II neuroblasts (middle, orange) divide into one self-renewing type II neuroblast and one intermediate neural precursor (INP). The INP starts expressing the neuroblast markers Asense and Deadpan to become a mature INP which divides asymmetrically into one GMC and one INP. In the optic lobe (bottom), neuroblasts arise from an epithelium. A wave of l(1)sc expression spreads through the epithelium and induces neuroblast formation. Optic lobe neuroblasts (blue) behave like type I neuroblasts although their precise lineage has not been determined. Differential expression of the markers Deadpan, Asense, Prospero and ElaV allows the unique identification of Individual cell types in type I, type II and optic lobe neuroblast lineages (see suppl. Fig. 1). b ∣ In Drosophila neuroblasts, the apically localized Par-3, Par-6, aPKC complex is connected to the Pins, Gα, Mud complex by the adaptor protein Inscuteable (Insc). During mitosis, this apical complex directs both the orientation of the mitotic spindle and the asymmetric localization of the adaptor proteins Pon and Miranda and the cell fate determinants Numb, Prospero and Brat to the opposite, basal cell cortex. After mitosis, Numb, Prospero and Brat act together to prevent self-renewal and induce cell cycle exit and differentiation. c ∣ In the C. elegans zygote, the anterior Par proteins Par-3, Par-6 and PKC-3 segregate into the anterior AB and the posterior Par-proteins Par-2 and Par-1 into the posterior P1 cell. Polarization starts after fertilization, when interactions between sperm centrosome and cortex allow Par-2 to accumulate. This initiates an anterior contraction of the cortical actin cytoskeleton, which allows anterior movement of Par-3 and Par-6.
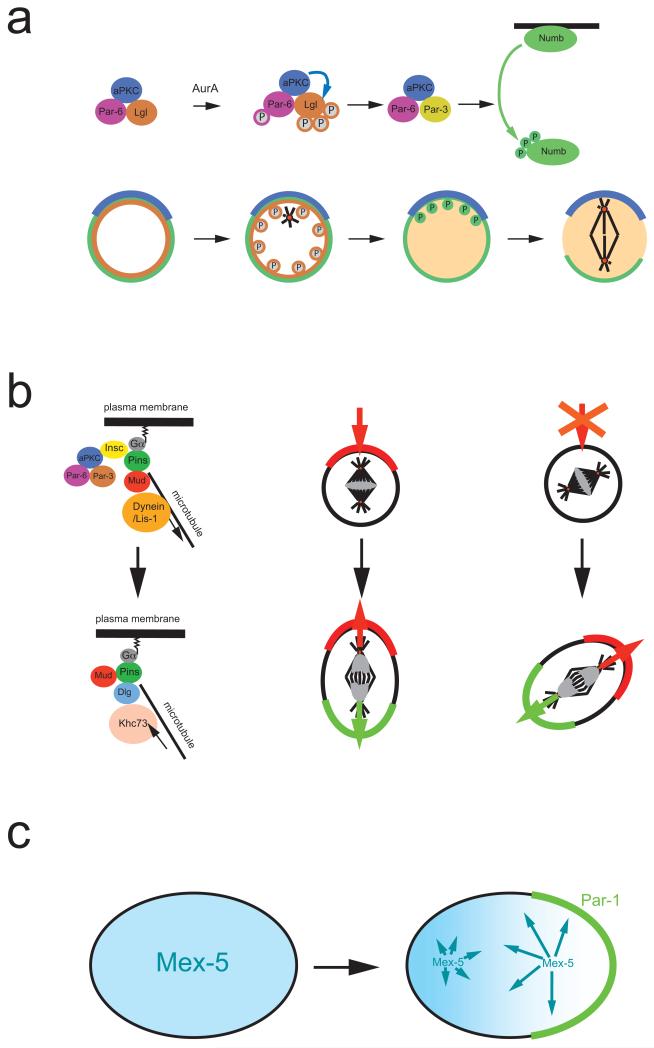
a ∣ In Drosophila melanogaster neuroblasts, activation of the kinase Aurora A triggers a subunit exchange in the apically localized Par-3–aPKC complex. Aurora A phosphorylates Par-6, which in turn activates aPKC, leading to Lgl phosphorylation and exit from the complex. Lgl is exchanged for Par-3, which acts as an adaptor that allows aPKC to phosphorylate of Numb. Phosphorylated Numb is released into the cytoplasm. Since aPKC is restricted to the apical cortex, Numb is retained on the basal side and segregates into the basal daughter cell. b ∣ Left side: In metaphase, Gα, Pins and Mud establish a cortical attachment site for astral microtubules to orient the mitotic spindle. In telophase, however, it is the mitotic spindle that influences cortical polarity via a pathway involving the kinesin Khc-73 and the protein Dlg. Right side: Normally, the telophase pathway is not essential. When components of the apical complex are missing, however, it will rescue the formation of opposing cortical domains in ana and telophase. The new polarity axis aligns with the mitotic spindle and no longer always with apical-basal polarity. c ∣ In C. elegans, Mex-5 (blue) and Pie-1 (not shown) exist as fast and slow diffusing forms. The faster diffusing form of Mex-5 is more abundant posteriorly while more rapidly diffusable Pie-1 is anterior, resulting in the asymmetric distribution of the cytoplasmic proteins. For Mex-5, phosphorylation by posteriorly localized Par-1 may be responsible for the faster diffusion rate.
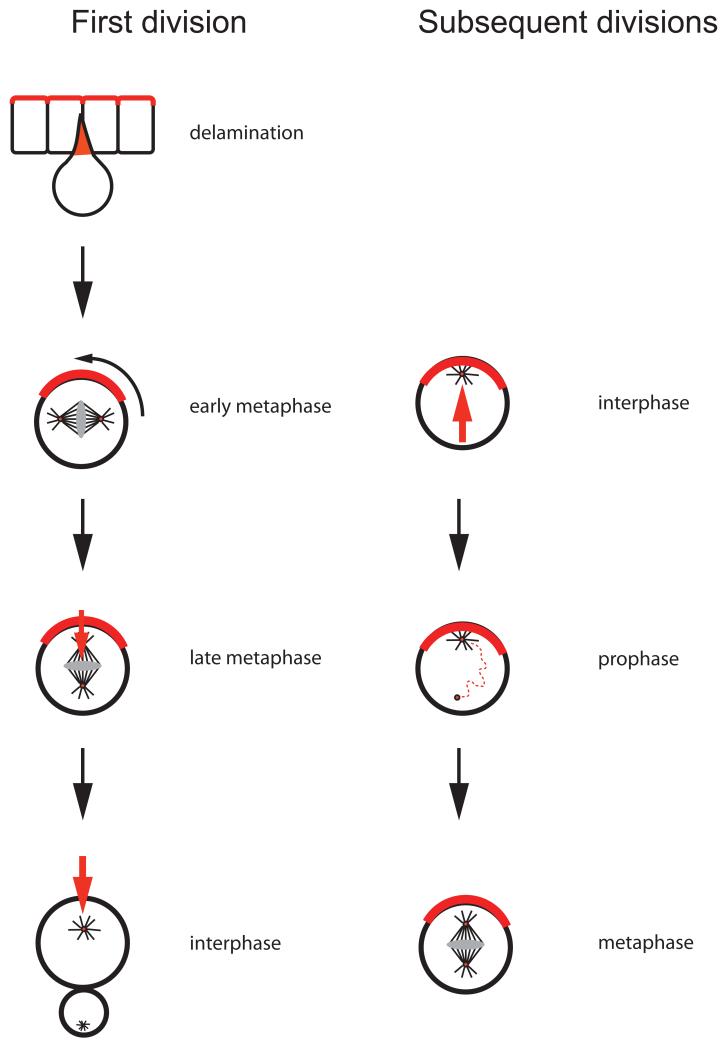
During the first neuroblast cycle, Par proteins are inherited from the apical cortex of the overlying epithelium. Through a series of adaptor proteins, they recruit Mud (red), which forms cortical attachment sites for astral microtubules and thereby induces spindle rotation into an apical–basal orientation. During subsequent cell cycles, centrosomes are responsible for maintaining cortical polarity in interphase. Cortical Pins, in turn, is required for maintaining the apical centrosome while the sister centriole migrates through the cytoplasm. On the basal side, this centriole recruits pericentriolar material to form a bipolar spindle in the proper apical-basal orientation.
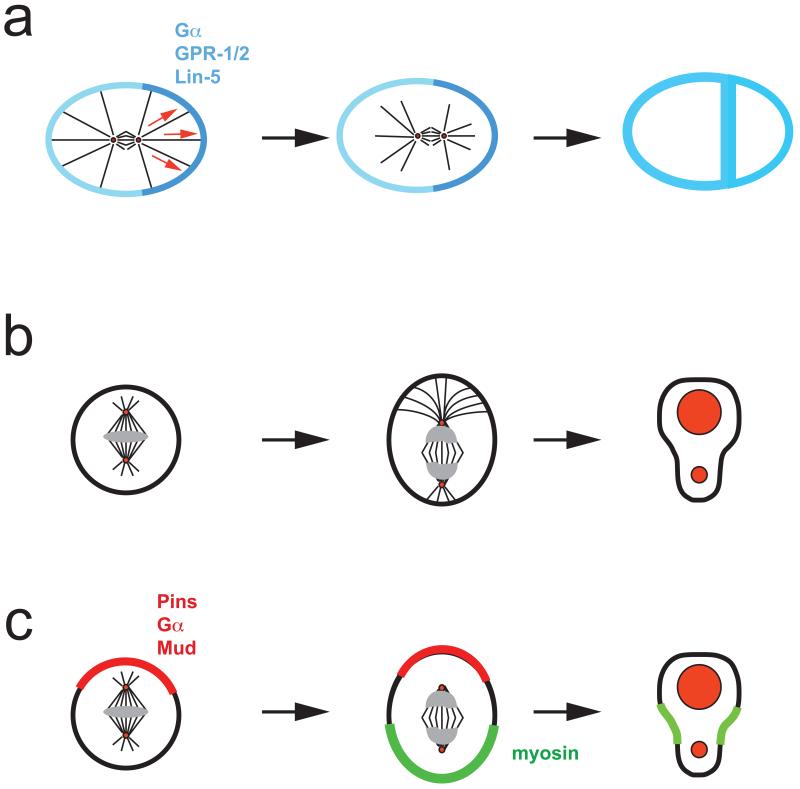
a ∣ In the C. elegans zygote, heterotrimeric G-protein α-subunits and the GoLoco proteins GPR-1 and GPR-2 recruit the dynein binding protein Lin-5 to the cortex to facilitate microtubule-cell cortex interactions. Higher concentration of GPR-1/2 on the posterior side, results in a net pulling force and posterior spindle displacement. b ∣ In Drosophila neuroblasts, the apical microtubule aster is larger resulting in spindle displacement towards the basal and asymmetric cleavage. c ∣ In neuroblasts, the apical Pins–Gα–Mud complex (red) induces a basal shift of cortical myosin,(green) resulting in basal displacement of the cleavage furrow. This pathway can generate asymmetric daughter cell sizes even in the absence of a mitotic spindle.
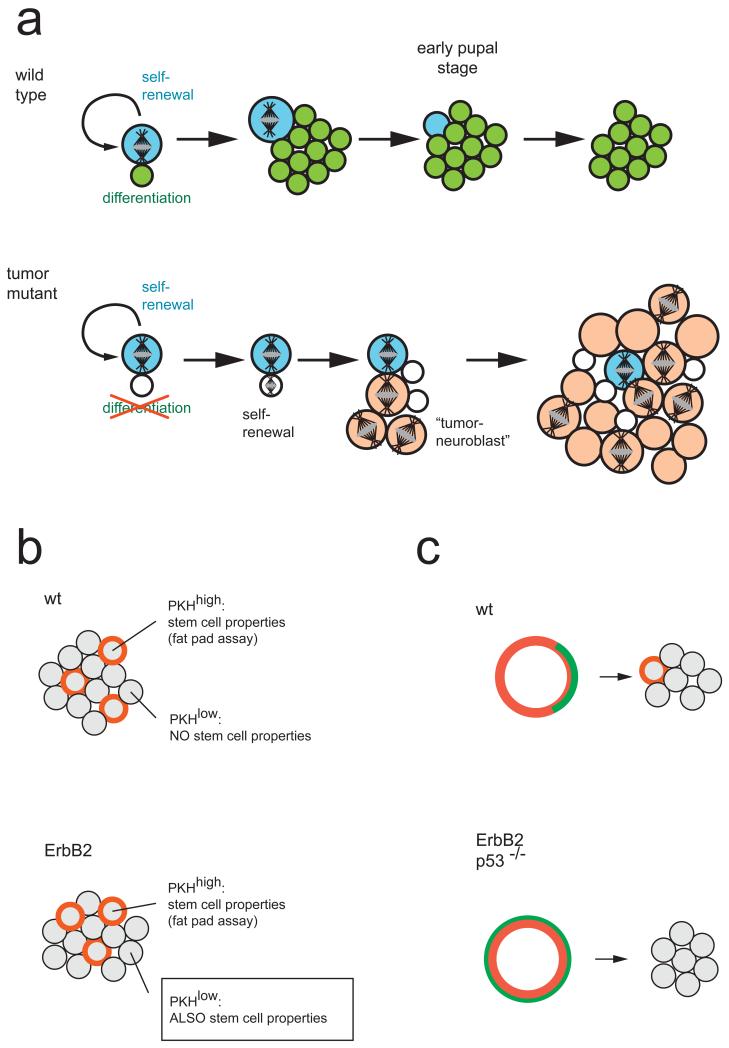
a ∣ Wild-type Drosophila neuroblasts generate one small self renewing (blue) and one large differentiating (green) daughter cell. The differentiating daughter cell exits the cell cycle after a terminal division (not shown). The neuroblast shrinks during pupal stages and undergoes apoptosis. In mutants affecting asymmetric cell division, the smaller daughter cell (white) cannot differentiate. After some time, it undergoes mitosis and reverts back to a tumour neuroblast (orange). These tumuor neuroblasts are abnormal because they do not exit the cell cycle during pupal stages. Whether or not the original neuroblast (blue) disappears or also continues to proliferate is unclear. b ∣ Mammospheres grown from wild-type mammary gland tissue or ErbB2 mutant tumour tissue contain the same number of slowly proliferating (PKH-26 dye (red) retaining) cells. In wild-type tissue, only the cells retaining the dye can form secondary mammospheres, but in tumour tissue, mammospheres can be grown from any cell. c ∣ PKH-26 dye-retaining cells can be purified by FACS sorting. In wild-type tissue, these cells localize Numb asymmetrically (green). When cultured, one dye-retaining cell remains, indicating that the initial division was asymmetric. When isolated from a ErbB2 tumour model or p53-mutant mice, PKH-26high cells do not divide asymmetrically and all daughter cells lose the dye, indicating that the initial division was symmetric.
References
-
- Conklin EG. The organization and cell-lineage of the ascidian egg. J. Acad. Nat. Sci. Philadelphia. 1905;13:1–119.
-
- Rhyu MS, Jan LY, Jan YN. Asymmetric distribution of numb protein during division of the sensory organ precursor cell confers distinct fates to daughter cells. Cell. 1994;76:477–491. - PubMed
-
- Spana EP, Kopczynski C, Goodman CS, Doe CQ. Asymmetric localization of numb autonomously determines sibling neuron identity in the Drosophila CNS. Development. 1995;121:3489–3494. - PubMed
-
- Uemura T, Shepherd S, Ackerman L, Jan LY, Jan YN. numb, a gene required in determination of cell fate during sensory organ formation in Drosophila embryos. Cell. 1989;58:349–360. - PubMed
-
- Kemphues KJ, Priess JR, Morton DG, Cheng NS. Identification of genes required for cytoplasmic localization in early C. elegans embryos. Cell. 1988;52:311–320. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
