

Potentiation of the glutamatergic synaptic input to rat locus coeruleus neurons by P2X7 receptors
- PMID: 21103218
- PMCID: PMC2947656
- DOI: 10.1007/s11302-010-9198-3
Potentiation of the glutamatergic synaptic input to rat locus coeruleus neurons by P2X7 receptors
Abstract
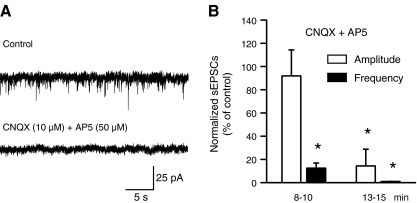
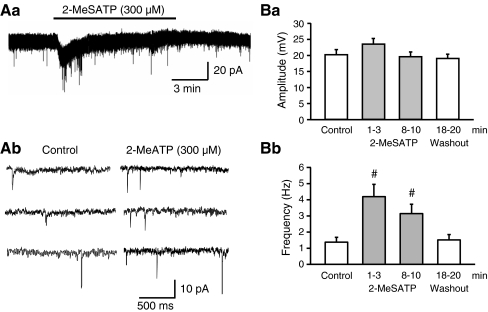
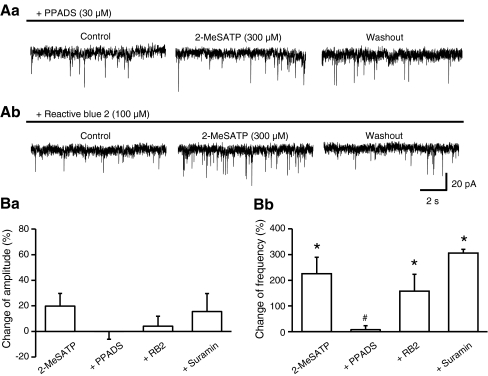
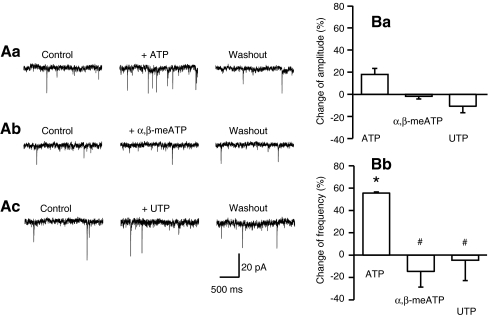
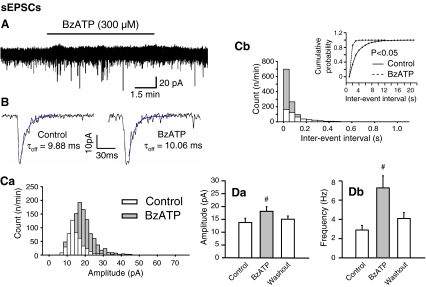
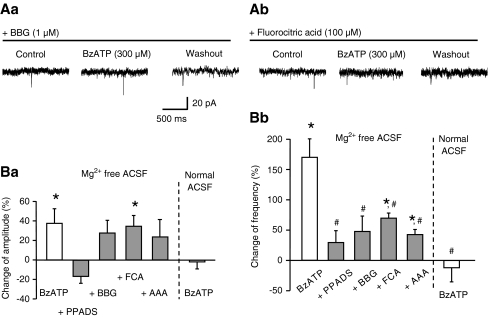
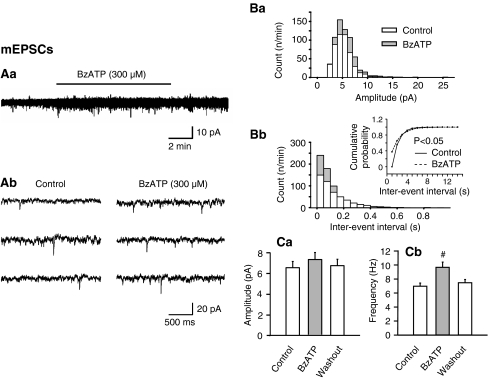
Locus coeruleus (LC) neurons in a rat brain slice preparation were superfused with a Mg(2+)-free and bicuculline-containing external medium. Under these conditions, glutamatergic spontaneous excitatory postsynaptic currents (sEPSCs) were recorded by means of the whole-cell patch-clamp method. ATP, as well as its structural analogue 2-methylthio ATP (2-MeSATP), both caused transient inward currents, which were outlasted by an increase in the frequency but not the amplitude of the sEPSCs. PPADS, but not suramin or reactive blue 2 counteracted both effects of 2-MeSATP. By contrast, α,β-methylene ATP (α,β-meATP), UTP and BzATP did not cause an inward current response. Of these latter agonists, only BzATP slightly facilitated the sEPSC amplitude and strongly potentiated its frequency. PPADS and Brilliant Blue G, as well as fluorocitric acid and aminoadipic acid prevented the activity of BzATP. Furthermore, BzATP caused a similar facilitation of the miniature (m)EPSC (recorded in the presence of tetrodotoxin) and sEPSC frequencies (recorded in its absence). Eventually, capsaicin augmented the frequency of the sEPSCs in a capsazepine-, but not PPADS-antagonizable, manner. In conclusion, the stimulation of astrocytic P2X7 receptors appears to lead to the outflow of a signalling molecule, which presynaptically increases the spontaneous release of glutamate onto LC neurons from their afferent fibre tracts. It is suggested, that the two algogenic compounds ATP and capsaicin utilise separate receptor systems to potentiate the release of glutamate and in consequence to increase the excitability of LC neurons.
Keywords: Adenosine 5′-triphosphate; Locus coeruleus; Miniature excitatory postsynaptic currents; P2X7 receptors; Presynaptic modulation; Spontaneous excitatory postsynaptic currents.
Figures

Similar articles
-
Supersensitivity of P2X receptors in cerebrocortical cell cultures after in vitro ischemia.J Neurochem. 2005 Dec;95(5):1421-37. doi: 10.1111/j.1471-4159.2005.03465.x. J Neurochem. 2005. PMID: 16313518
-
Role of ATP in fast excitatory synaptic potentials in locus coeruleus neurones of the rat.Br J Pharmacol. 1997 Oct;122(3):423-30. doi: 10.1038/sj.bjp.0701386. Br J Pharmacol. 1997. PMID: 9351497 Free PMC article.
-
Excitation of rat spinal ventral horn neurons by purinergic P2X and P2Y receptor activation.Brain Res. 2010 Jun 22;1340:10-7. doi: 10.1016/j.brainres.2010.04.053. Epub 2010 Apr 25. Brain Res. 2010. PMID: 20423703
-
P2X7-like receptor subunits enhance excitatory synaptic transmission at central synapses by presynaptic mechanisms.Neuroscience. 2004;128(2):269-80. doi: 10.1016/j.neuroscience.2004.06.014. Neuroscience. 2004. PMID: 15350640
-
Modulation of allopregnanolone on excitatory transmitters release from single glutamatergic terminal.Brain Res Bull. 2013 Apr;93:39-46. doi: 10.1016/j.brainresbull.2012.11.002. Epub 2012 Nov 19. Brain Res Bull. 2013. PMID: 23174309 Review.
Cited by
-
Neurodevelopmental alterations and seizures developed by mouse model of infantile hypophosphatasia are associated with purinergic signalling deregulation.Hum Mol Genet. 2016 Oct 1;25(19):4143-4156. doi: 10.1093/hmg/ddw248. Epub 2016 Jul 27. Hum Mol Genet. 2016. PMID: 27466191 Free PMC article.
-
Neuronal P2X7 Receptors Revisited: Do They Really Exist?J Neurosci. 2017 Jul 26;37(30):7049-7062. doi: 10.1523/JNEUROSCI.3103-16.2017. J Neurosci. 2017. PMID: 28747388 Free PMC article. Review.
-
Modulation of excitatory neurotransmission by neuronal/glial signalling molecules: interplay between purinergic and glutamatergic systems.Purinergic Signal. 2016 Mar;12(1):1-24. doi: 10.1007/s11302-015-9480-5. Epub 2015 Nov 6. Purinergic Signal. 2016. PMID: 26542977 Free PMC article. Review.
-
Effect of capsaicin on thermoregulation: an update with new aspects.Temperature (Austin). 2015 Jun 2;2(2):277-96. doi: 10.1080/23328940.2015.1048928. eCollection 2015 Apr-Jun. Temperature (Austin). 2015. PMID: 27227029 Free PMC article. Review.
-
Pathophysiology of astroglial purinergic signalling.Purinergic Signal. 2012 Sep;8(3):629-57. doi: 10.1007/s11302-012-9300-0. Epub 2012 May 1. Purinergic Signal. 2012. PMID: 22544529 Free PMC article. Review.
References
-
- Stamford JA. Descending control of pain. Br J Anaesth. 1995;75:217–227. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
