

Biliverdin inhibits hepatitis C virus nonstructural 3/4A protease activity: mechanism for the antiviral effects of heme oxygenase?
- PMID: 21105106
- PMCID: PMC3058505
- DOI: 10.1002/hep.23921
Biliverdin inhibits hepatitis C virus nonstructural 3/4A protease activity: mechanism for the antiviral effects of heme oxygenase?
Abstract
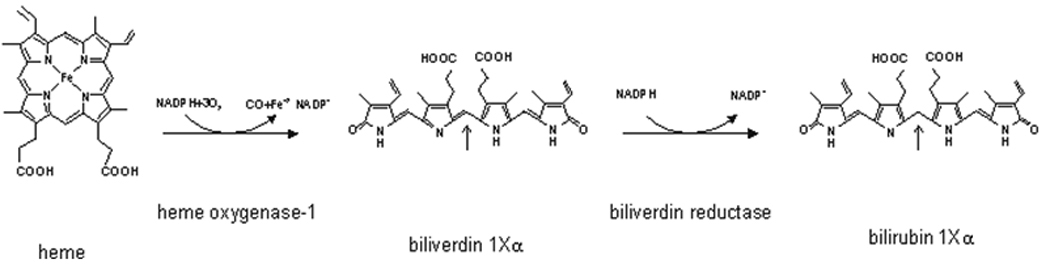
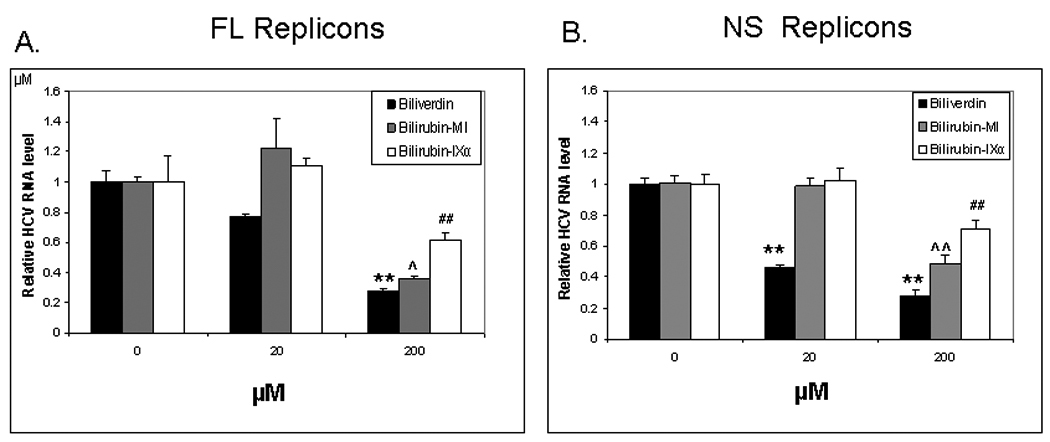
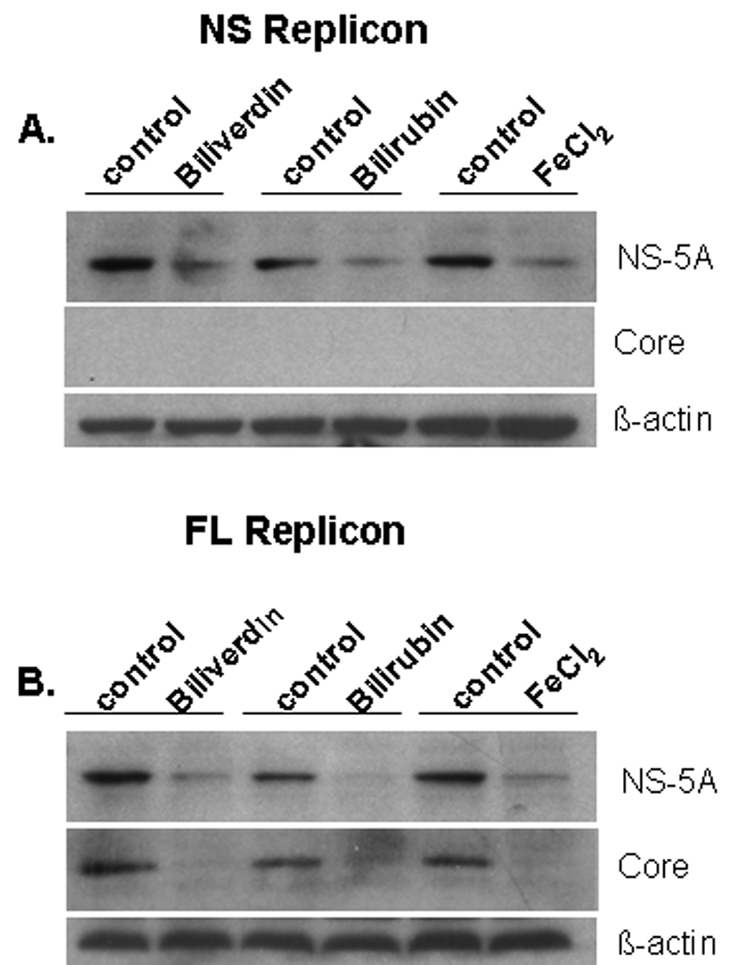
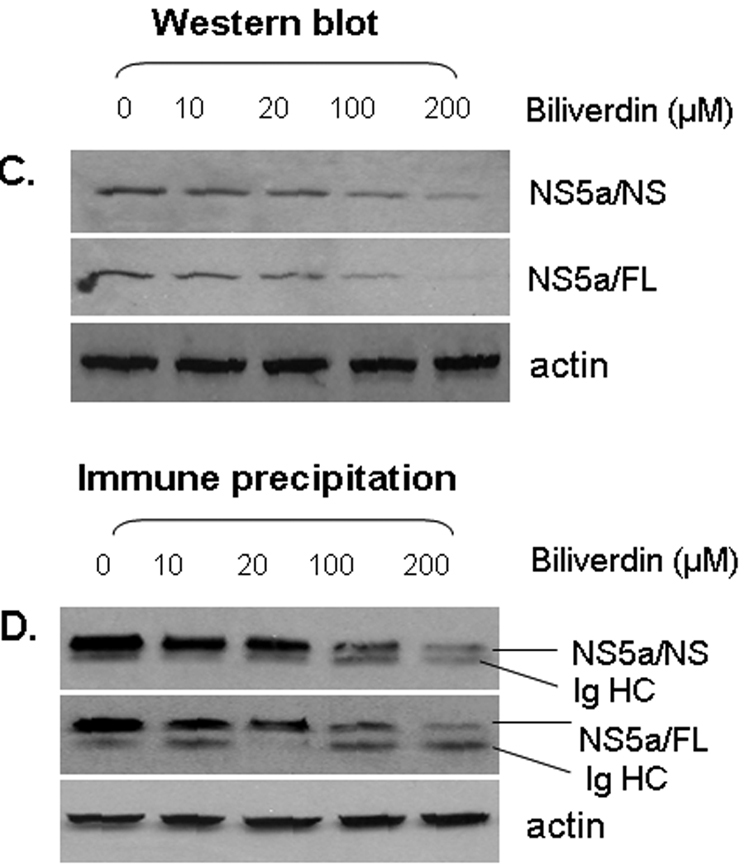
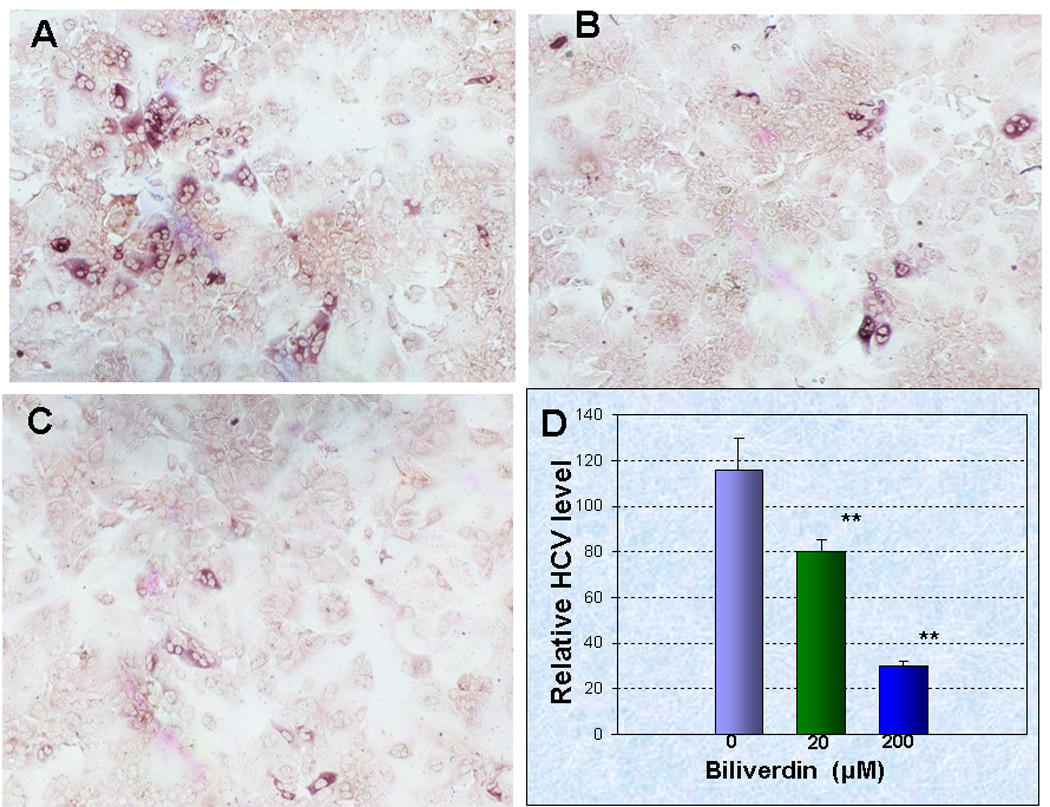
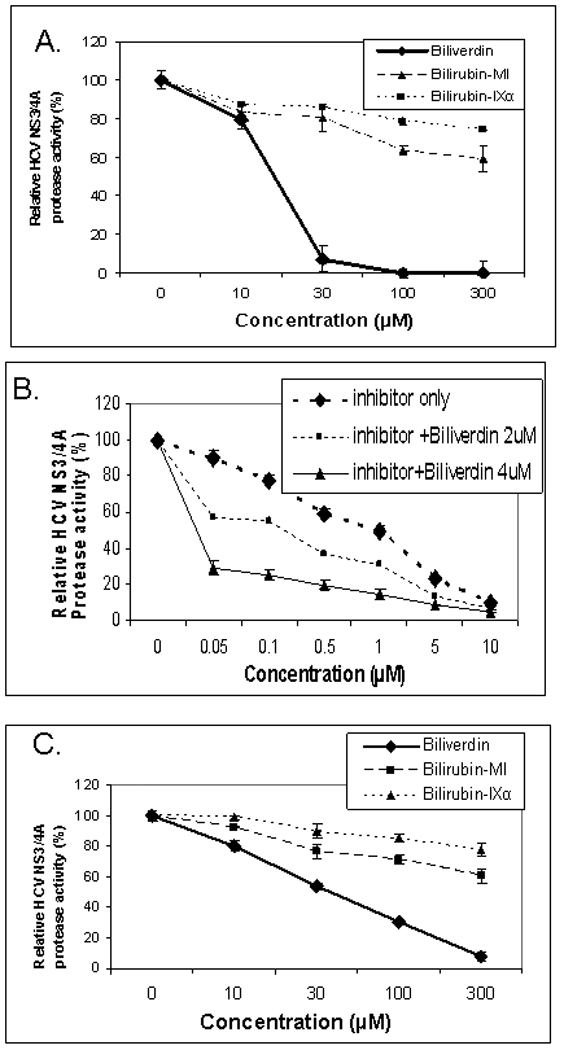
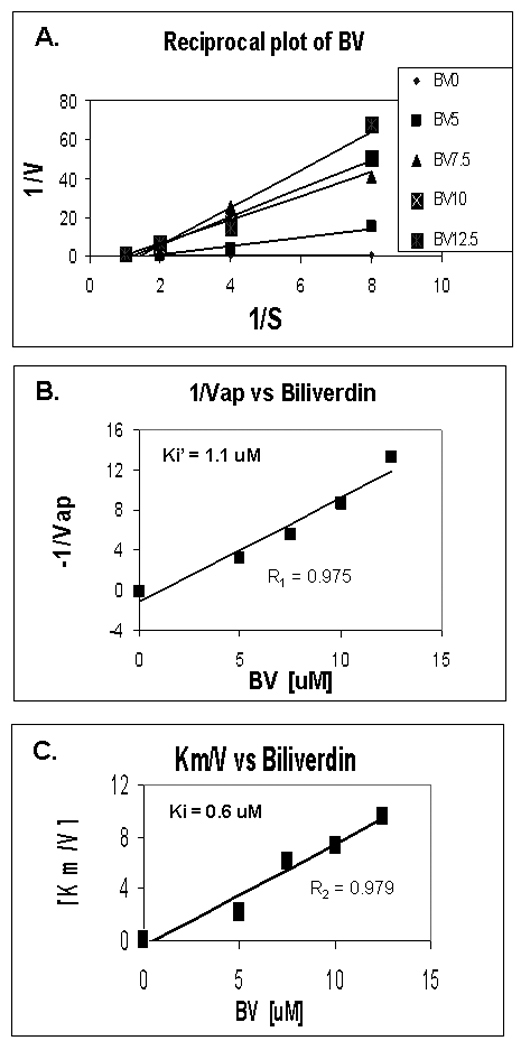
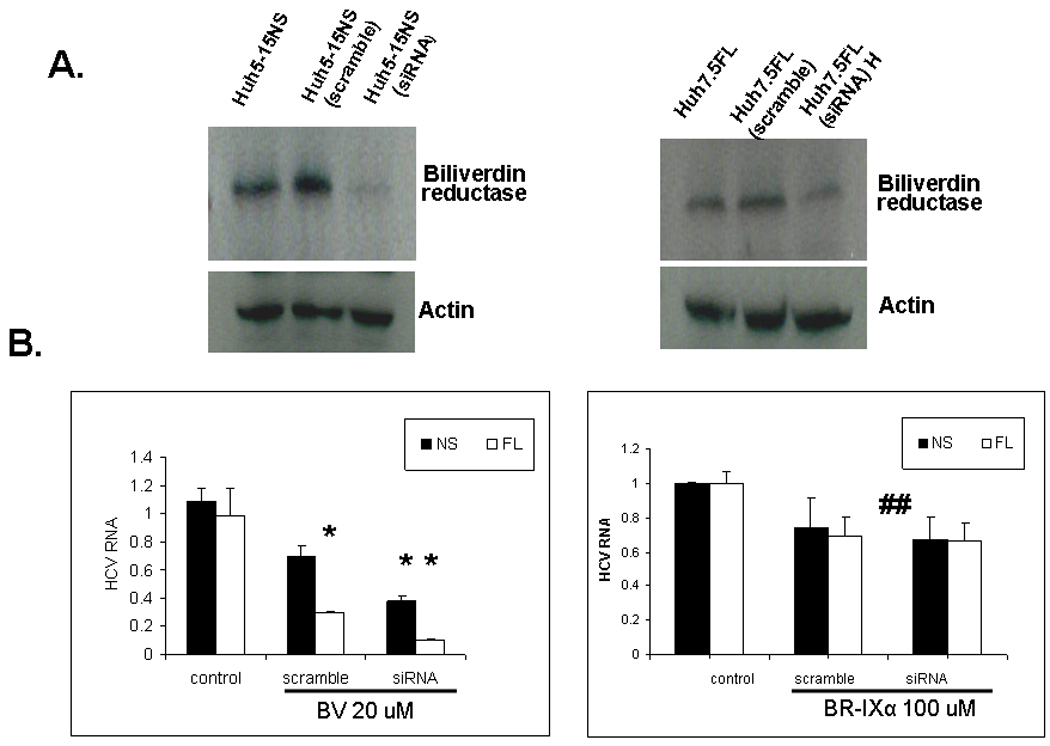
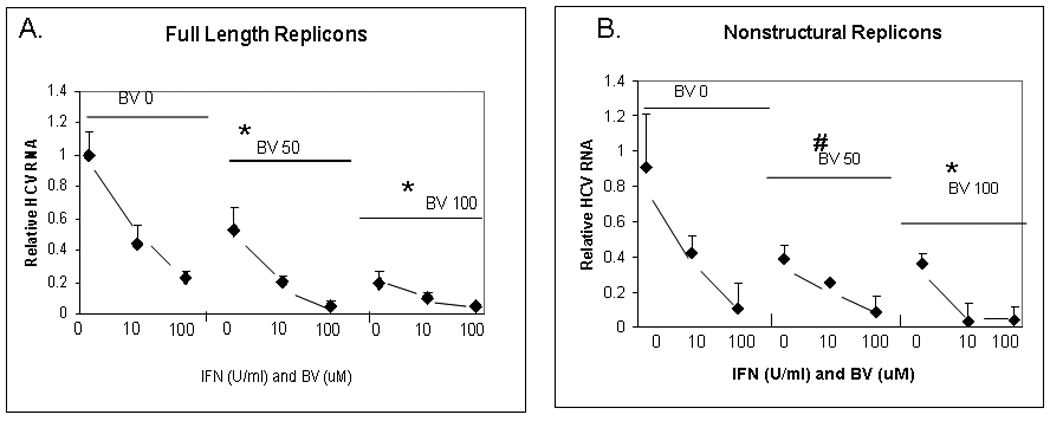
Induction of heme oxygenase-1 (HO-1) inhibits hepatitis C virus (HCV) replication. Of the products of the reaction catalyzed by HO-1, iron has been shown to inhibit HCV ribonucleic acid (RNA) polymerase, but little is known about the antiviral activity of biliverdin (BV). Herein, we report that BV inhibits viral replication and viral protein expression in a dose-dependent manner in replicons and cells harboring the infectious J6/JFH construct. Using the SensoLyte 620 HCV Protease Assay with a wide wavelength excitation/emission (591 nm/622 nm) fluorescence energy transfer peptide, we found that both recombinant and endogenous nonstructural 3/4A (NS3/4A) protease from replicon microsomes are potently inhibited by BV. Of the tetrapyrroles tested, BV was the strongest inhibitor of NS3/4A activity, with a median inhibitory concentration (IC(50)) of 9 μM, similar to that of the commercial inhibitor, AnaSpec (Fremont, CA) #25346 (IC(50) 5 μM). Lineweaver-Burk plots indicated mixed competitive and noncompetitive inhibition of the protease by BV. In contrast, the effects of bilirubin (BR) on HCV replication and NS3/4A were much less potent. Because BV is rapidly converted to BR by biliverdin reductase (BVR) intracellularly, the effect of BVR knockdown on BV antiviral activity was assessed. After greater than 80% silencing of BVR, inhibition of viral replication by BV was enhanced. BV also increased the antiviral activity of α-interferon in replicons.
Conclusion: BV is a potent inhibitor of HCV NS3/4A protease, which likely contributes to the antiviral activity of HO-1. These findings suggest that BV or its derivatives may be useful in future drug therapies targeting the NS3/4A protease.
Copyright © 2010 American Association for the Study of Liver Diseases.
Conflict of interest statement
None of the authors who participated in this study have commercial or other associations that might pose a conflict of interest.
Figures
Comment in
-
Biliverdin and heme oxygenase antiviral activity against hepatitis C virus.Ann Hepatol. 2011 Jan-Mar;10(1):105-7. Ann Hepatol. 2011. PMID: 21301021 No abstract available.
Similar articles
-
Restoration of type I interferon expression by heme and related tetrapyrroles through inhibition of NS3/4A protease.J Infect Dis. 2013 Nov 15;208(10):1653-63. doi: 10.1093/infdis/jit338. Epub 2013 Jul 29. J Infect Dis. 2013. PMID: 23901085 Free PMC article.
-
Lucidone suppresses hepatitis C virus replication by Nrf2-mediated heme oxygenase-1 induction.Antimicrob Agents Chemother. 2013 Mar;57(3):1180-91. doi: 10.1128/AAC.02053-12. Epub 2012 Dec 17. Antimicrob Agents Chemother. 2013. PMID: 23254429 Free PMC article.
-
Inhibition of protease-inhibitor-resistant hepatitis C virus replicons and infectious virus by intracellular intrabodies.Antiviral Res. 2010 Oct;88(1):95-106. doi: 10.1016/j.antiviral.2010.08.001. Epub 2010 Aug 10. Antiviral Res. 2010. PMID: 20705106 Free PMC article.
-
Discovery and development of VX-950, a novel, covalent, and reversible inhibitor of hepatitis C virus NS3.4A serine protease.Infect Disord Drug Targets. 2006 Mar;6(1):3-16. doi: 10.2174/187152606776056706. Infect Disord Drug Targets. 2006. PMID: 16787300 Review.
-
Hepatitis C virus NS3-4A protease inhibitors: countering viral subversion in vitro and showing promise in the clinic.Curr Opin Drug Discov Devel. 2006 Sep;9(5):606-17. Curr Opin Drug Discov Devel. 2006. PMID: 17002221 Review.
Cited by
-
Mesobiliverdin IXα Enhances Rat Pancreatic Islet Yield and Function.Front Pharmacol. 2013 Apr 23;4:50. doi: 10.3389/fphar.2013.00050. eCollection 2013. Front Pharmacol. 2013. PMID: 23630498 Free PMC article.
-
Dengue Virus Targets Nrf2 for NS2B3-Mediated Degradation Leading to Enhanced Oxidative Stress and Viral Replication.J Virol. 2020 Nov 23;94(24):e01551-20. doi: 10.1128/JVI.01551-20. Print 2020 Nov 23. J Virol. 2020. PMID: 32999020 Free PMC article.
-
Multiple origins of green blood in New Guinea lizards.Sci Adv. 2018 May 16;4(5):eaao5017. doi: 10.1126/sciadv.aao5017. eCollection 2018 May. Sci Adv. 2018. PMID: 29774232 Free PMC article.
-
Ribavirin induced hemolysis: a novel mechanism of action against chronic hepatitis C virus infection.World J Gastroenterol. 2014 Nov 21;20(43):16184-90. doi: 10.3748/wjg.v20.i43.16184. World J Gastroenterol. 2014. PMID: 25473172 Free PMC article. Review.
-
Short communication: atazanavir-based therapy is associated with higher hepatitis C viral load in HIV type 1-infected subjects with untreated hepatitis C.AIDS Res Hum Retroviruses. 2013 Feb;29(2):223-5. doi: 10.1089/AID.2012.0126. Epub 2012 Oct 10. AIDS Res Hum Retroviruses. 2013. PMID: 22966845 Free PMC article.
References
-
- Firpi RJ, Nelson DR. Current and future hepatitis C therapies. Archives of Medical Research. 2007 Aug;38(6):678–690. - PubMed
-
- Lindenbach BD, Rice CM. Unravelling hepatitis C virus replication from genome to function. Nature. 2005 Aug 18;436(7053):933–938. - PubMed
-
- Soriano V, Peters MG, Zeuzem S. New Therapies for Hepatitis C Virus Infection. Clinical Infectious Diseases. 2009 Feb 1;48(3):313–320. - PubMed
-
- Choi J, Ou JHJ. Mechanisms of liver injury. III. Oxidative stress in the pathogenesis of hepatitis C virus. American Journal of Physiology-Gastrointestinal and Liver Physiology. 2006 May;290(5):G847–G851. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources