

Post-transcriptional defects of antioxidant selenoenzymes cause oxidative stress under methylmercury exposure
- PMID: 21106535
- PMCID: PMC3057802
- DOI: 10.1074/jbc.M110.168872
Post-transcriptional defects of antioxidant selenoenzymes cause oxidative stress under methylmercury exposure
Abstract
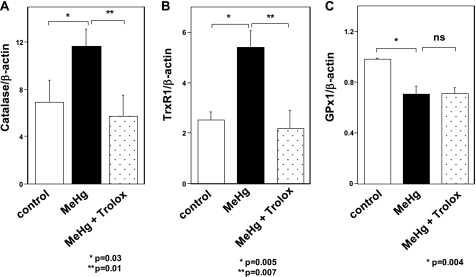
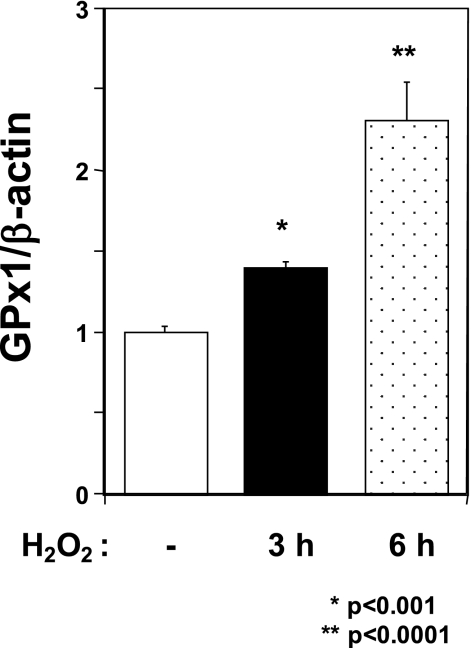
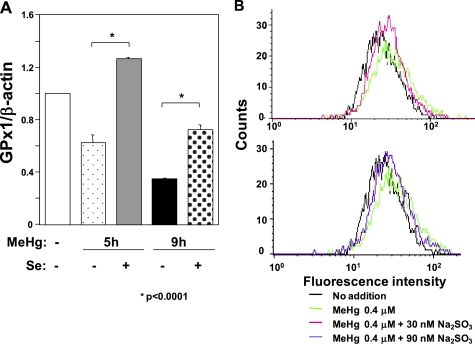
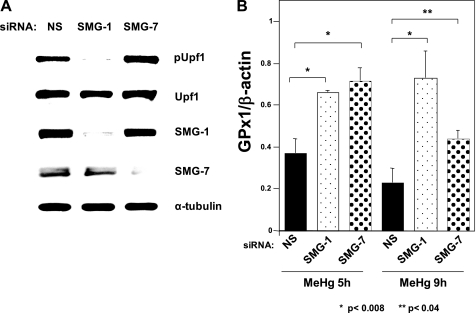
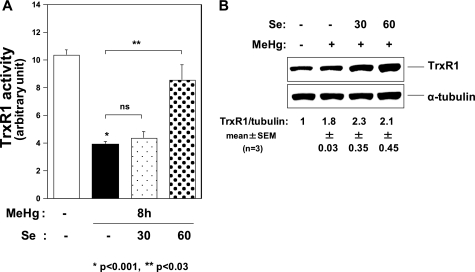
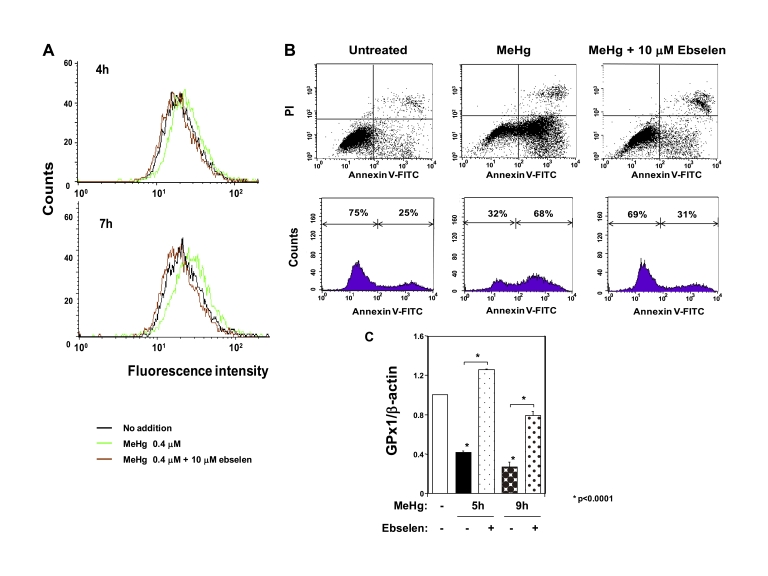
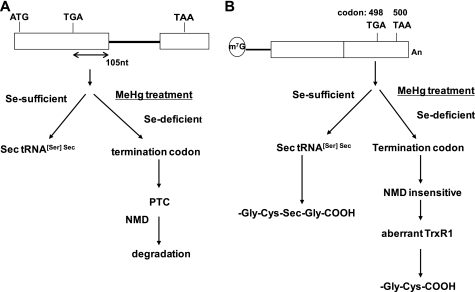
Methylmercury (MeHg) toxicity is a continuous environmental problem to human health. The critical role of oxidative stress in the pathogenesis of MeHg cytotoxicity has been clarified, but the molecular mechanisms underlying MeHg-mediated oxidative stress remain to be elucidated. Here we demonstrate a post-transcriptional effect of MeHg on antioxidant selenoenzymes by using a MeHg-susceptible cell line. MeHg-induced selenium deficiency leads to failure of the recoding of a UGA codon for selenocysteine and results in degradation of the major antioxidant selenoenzyme glutathione peroxidase 1 (GPx1) mRNA by nonsense-mediated mRNA decay (NMD), a cellular mechanism that detects the premature termination codon (PTC) located 5'-upstream of the last exon-exon junction and degrades PTC-containing mRNAs. In contrast, thioredoxin reductase 1 (TrxR1), another antioxidant selenoenzyme of the thioredoxin system, was likely skipped by NMD because of a UGA codon in the last exon. However, TrxR1 activity was decreased despite mRNA up-regulation, which was probably due to the synthesis of aberrant TrxR1 protein without selenocysteine. Changes in selenoenzyme GPx1 and TrxR1 mRNAs were observed earlier than was the incidence of oxidative stress and up-regulation of other antioxidant enzyme mRNAs. Results indicated that the MeHg-induced relative selenium-deficient condition affects the major antioxidant selenoenzymes GPx1 and TrxR1 through a post-transcriptional effect, resulting in the disturbance of cellular redox systems and the incidence of oxidative stress. Treatment with ebselen, a seleno-organic compound, effectively suppressed oxidative stress and protected cells against MeHg-induced relative selenium deficiency and cytotoxicity.
Figures


References
-
- Hamada R., Osame M. (1996) Toxicology of Metals (Chang L. W. ed) pp. 337–351, CRC Press, London
-
- Bakir F., Damluji S. F., Amin-Zaki L., Murtadha M., Khalidi A., al-Rawi N. Y., Tikriti S., Dahahir H. I., Clarkson T. W., Smith J. C., Doherty R. A. (1973) Science 181, 230–241 - PubMed
-
- Davis L. E., Kornfeld M., Mooney H. S., Fiedler K. J., Haaland K. Y., Orrison W. W., Cernichiari E., Clarkson T. W. (1994) Ann. Neurol. 35, 680–688 - PubMed
-
- Deleted in proof.
-
- Park S. T., Lim K. T., Chung Y. T., Kim S. U. (1996) Neurotoxicology 17, 37–45 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
