

Evolution of adaptive phenotypic variation patterns by direct selection for evolvability
- PMID: 21106581
- PMCID: PMC3097830
- DOI: 10.1098/rspb.2010.2113
Evolution of adaptive phenotypic variation patterns by direct selection for evolvability
Abstract
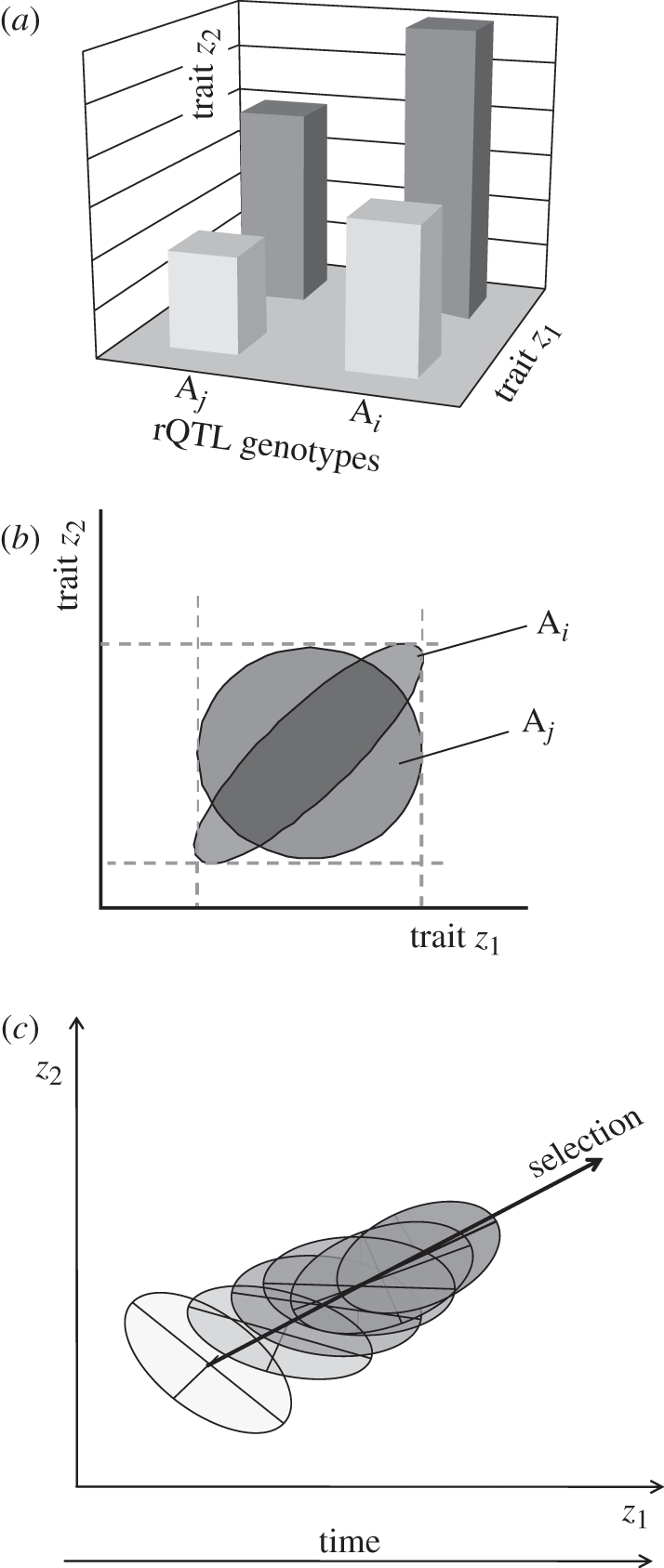
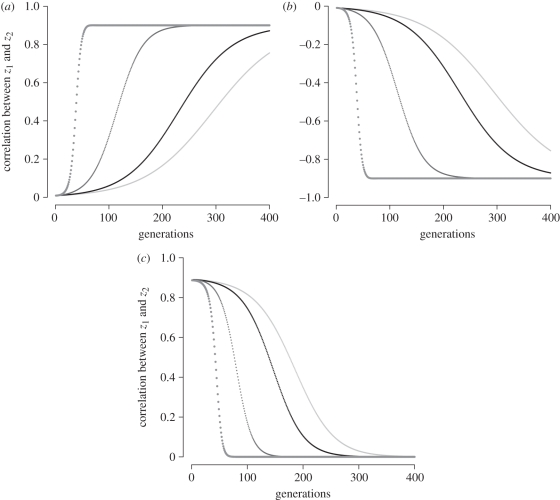
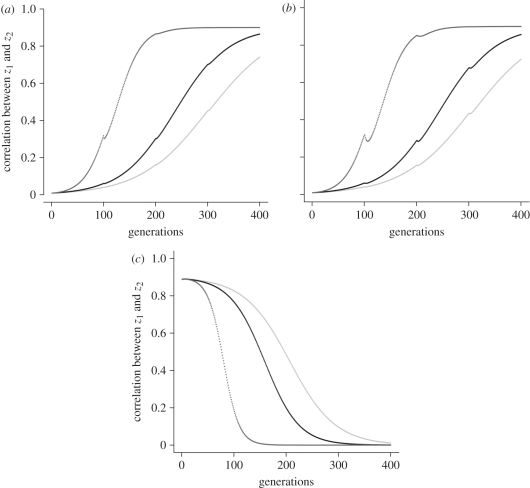
A basic assumption of the Darwinian theory of evolution is that heritable variation arises randomly. In this context, randomness means that mutations arise irrespective of the current adaptive needs imposed by the environment. It is broadly accepted, however, that phenotypic variation is not uniformly distributed among phenotypic traits, some traits tend to covary, while others vary independently, and again others barely vary at all. Furthermore, it is well established that patterns of trait variation differ among species. Specifically, traits that serve different functions tend to be less correlated, as for instance forelimbs and hind limbs in bats and humans, compared with the limbs of quadrupedal mammals. Recently, a novel class of genetic elements has been identified in mouse gene-mapping studies that modify correlations among quantitative traits. These loci are called relationship loci, or relationship Quantitative Trait Loci (rQTL), and affect trait correlations by changing the expression of the existing genetic variation through gene interaction. Here, we present a population genetic model of how natural selection acts on rQTL. Contrary to the usual neo-Darwinian theory, in this model, new heritable phenotypic variation is produced along the selected dimension in response to directional selection. The results predict that selection on rQTL leads to higher correlations among traits that are simultaneously under directional selection. On the other hand, traits that are not simultaneously under directional selection are predicted to evolve lower correlations. These results and the previously demonstrated existence of rQTL variation, show a mechanism by which natural selection can directly enhance the evolvability of complex organisms along lines of adaptive change.
Figures
Similar articles
-
The mutation matrix and the evolution of evolvability.Evolution. 2007 Apr;61(4):727-45. doi: 10.1111/j.1558-5646.2007.00071.x. Evolution. 2007. PMID: 17439608
-
Phenotypic plasticity facilitates mutational variance, genetic variance, and evolvability along the major axis of environmental variation.Evolution. 2012 Sep;66(9):2891-902. doi: 10.1111/j.1558-5646.2012.01649.x. Epub 2012 May 1. Evolution. 2012. PMID: 22946810
-
Selection at linked sites shapes heritable phenotypic variation in C. elegans.Science. 2010 Oct 15;330(6002):372-6. doi: 10.1126/science.1194208. Science. 2010. PMID: 20947766 Free PMC article.
-
The relationship between development and evolution through heritable variation.Novartis Found Symp. 2007;284:55-65; discussion 65-70, 110-5. doi: 10.1002/9780470319390.ch4. Novartis Found Symp. 2007. PMID: 17710847 Review.
-
Variation, selection and evolution of function-valued traits.Genetica. 2001;112-113:87-104. Genetica. 2001. PMID: 11838789 Review.
Cited by
-
Developmental bias as a cause and consequence of adaptive radiation and divergence.Front Cell Dev Biol. 2024 Oct 16;12:1453566. doi: 10.3389/fcell.2024.1453566. eCollection 2024. Front Cell Dev Biol. 2024. PMID: 39479512 Free PMC article. Review.
-
The more g-loaded, the more heritable, evolvable, and phenotypically variable: Homology with humans in chimpanzee cognitive abilities.Intelligence. 2015 May-Jun;50:159-163. doi: 10.1016/j.intell.2015.04.002. Intelligence. 2015. PMID: 26005227 Free PMC article.
-
The evolution of genetic covariance and modularity as a result of multigenerational environmental fluctuation.Evol Lett. 2023 Oct 17;7(6):457-466. doi: 10.1093/evlett/qrad048. eCollection 2023 Dec. Evol Lett. 2023. PMID: 38045721 Free PMC article.
-
Evolutionary Connectionism: Algorithmic Principles Underlying the Evolution of Biological Organisation in Evo-Devo, Evo-Eco and Evolutionary Transitions.Evol Biol. 2016;43(4):553-581. doi: 10.1007/s11692-015-9358-z. Epub 2015 Dec 8. Evol Biol. 2016. PMID: 27932852 Free PMC article.
-
Developmental and genetic origins of murine long bone length variation.J Exp Zool B Mol Dev Evol. 2011 Mar 15;316B(2):146-61. doi: 10.1002/jez.b.21388. Epub 2010 Dec 1. J Exp Zool B Mol Dev Evol. 2011. PMID: 21328530 Free PMC article.
References
-
- Terentjev P. V. 1931. Biometrische Untersuchungen ueber die morphologischen Merkmale von Rana ridibunda Pall. (Amphibia, Salientia). Biometrika 23, 23–51
-
- Berg R. L. 1960. The ecological significance of correlation pleiades. Evolution 14, 171–18010.2307/2405824 (doi:10.2307/2405824) - DOI - DOI
-
- Chetverikov S. S., Barker M., Lerner I. M. 1961. On certain aspects of the evolutionary process from the standpoint of modern genetics. Proc. Am. Phil. Soc. 105, 167–195
-
- Darwin C. 1859. On the origin of species. London, UK: John Murray
-
- Olson E. C., Miller R. L. 1958. Morphological integration. Chicago, IL: University of Chicago Press
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
