

Role of the CM2 protein in the influenza C virus replication cycle
- PMID: 21106743
- PMCID: PMC3020500
- DOI: 10.1128/JVI.01367-10
Role of the CM2 protein in the influenza C virus replication cycle
Abstract
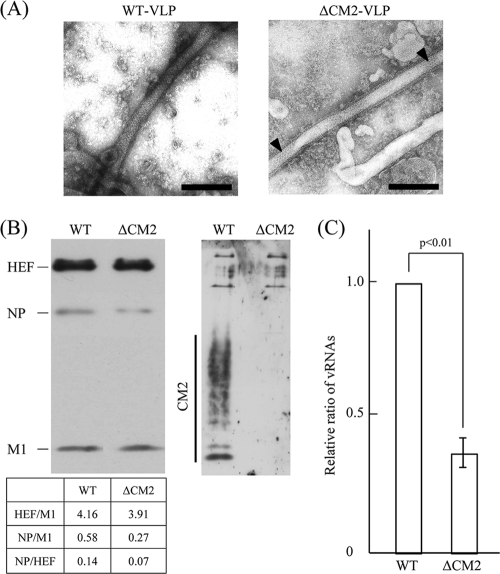
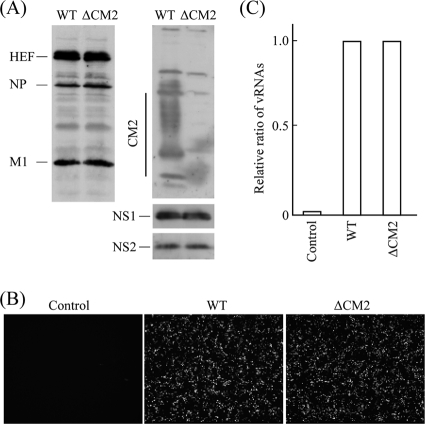
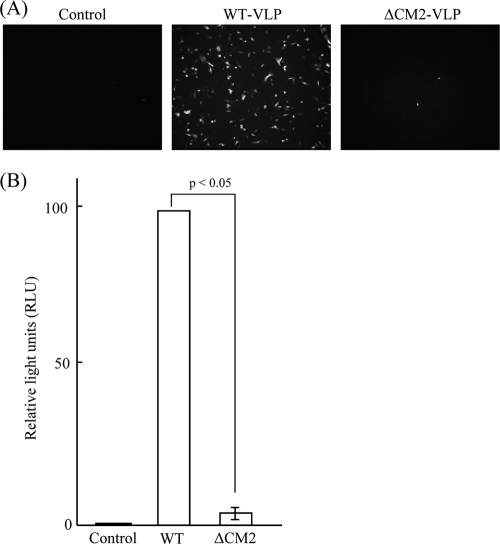
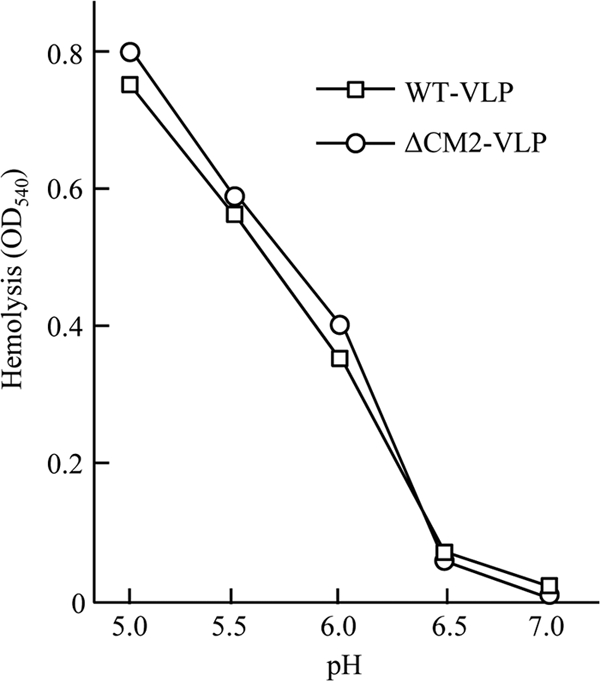
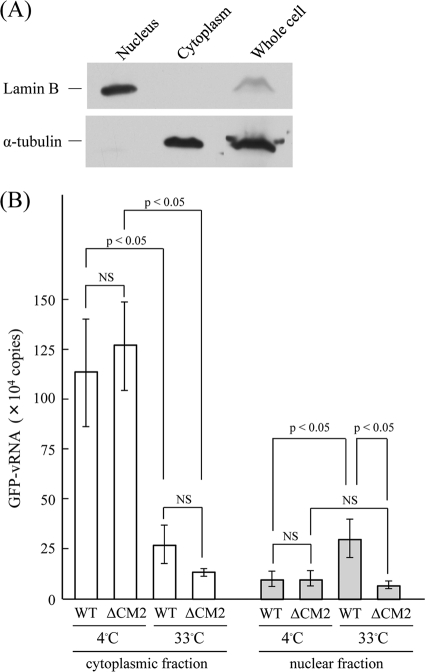
CM2 is the second membrane protein of influenza C virus. Although its biochemical characteristics, coding strategy, and properties as an ion channel have been extensively studied, the role(s) of CM2 in the virus replication cycle remains to be clarified. In order to elucidate this role, in the present study we generated CM2-deficient influenza C virus-like particles (VLPs) and examined the VLP-producing 293T cells, VLPs, and VLP-infected HMV-II cells. Quantification of viral RNA (vRNA) in the VLPs by real-time PCR revealed that the CM2-deficient VLPs contain approximately one-third of the vRNA found in wild-type VLPs although no significant differences were detected in the expression levels of viral components in VLP-producing cells or in the number and morphology of the generated VLPs. This finding suggests that CM2 is involved in the genome packaging process into VLPs. Furthermore, HMV-II cells infected with CM2-deficient VLPs exhibited significantly reduced reporter gene expression. Although CM2-deficient VLPs could be internalized into HMV-II cells as efficiently as wild-type VLPs, a smaller amount of vRNA was detected in the nuclear fraction of CM2-deficient VLP-infected cells than in that of wild-type VLP-infected cells, suggesting that the uncoating process of the CM2-deficient VLPs in the infected cells did not proceed in an appropriate manner. Taken together, the data obtained in the present study indicate that CM2 has a potential role in the genome packaging and uncoating processes of the virus replication cycle.
Figures
Similar articles
-
Glycosylation of CM2 is important for efficient replication of influenza C virus.Virology. 2012 Nov 10;433(1):167-75. doi: 10.1016/j.virol.2012.08.010. Epub 2012 Aug 22. Virology. 2012. PMID: 22921534
-
Effect of cysteine mutations in the extracellular domain of CM2 on the influenza C virus replication.PLoS One. 2013 Apr 4;8(4):e60510. doi: 10.1371/journal.pone.0060510. Print 2013. PLoS One. 2013. PMID: 23593230 Free PMC article.
-
Effect of Phosphorylation of CM2 Protein on Influenza C Virus Replication.J Virol. 2017 Oct 27;91(22):e00773-17. doi: 10.1128/JVI.00773-17. Print 2017 Nov 15. J Virol. 2017. PMID: 28878070 Free PMC article.
-
The molecular virology and reverse genetics of influenza C virus.Jpn J Infect Dis. 2010 May;63(3):157-65. Jpn J Infect Dis. 2010. PMID: 20495266 Review.
-
[Type C influenza].Nihon Rinsho. 2006 Oct;64(10):1942-9. Nihon Rinsho. 2006. PMID: 17037372 Review. Japanese.
Cited by
-
Prediction of Site Directed miRNAs as Key Players of Transcriptional Regulators Against Influenza C Virus Infection Through Computational Approaches.Front Mol Biosci. 2022 Apr 8;9:866072. doi: 10.3389/fmolb.2022.866072. eCollection 2022. Front Mol Biosci. 2022. PMID: 35463952 Free PMC article.
-
Temporal and Gene Reassortment Analysis of Influenza C Virus Outbreaks in Hong Kong, SAR, China.J Virol. 2022 Feb 9;96(3):e0192821. doi: 10.1128/JVI.01928-21. Epub 2021 Nov 17. J Virol. 2022. PMID: 34787455 Free PMC article.
-
Host Range, Biology, and Species Specificity of Seven-Segmented Influenza Viruses-A Comparative Review on Influenza C and D.Pathogens. 2021 Dec 5;10(12):1583. doi: 10.3390/pathogens10121583. Pathogens. 2021. PMID: 34959538 Free PMC article. Review.
-
The effect of the cytoplasmic tail of influenza C virus CM2 protein on its biochemical properties and intracellular processing.Biochem Biophys Rep. 2015 Jul 8;3:1-6. doi: 10.1016/j.bbrep.2015.07.001. eCollection 2015 Sep. Biochem Biophys Rep. 2015. PMID: 29124162 Free PMC article.
-
Viroporins in the Influenza Virus.Cells. 2019 Jun 29;8(7):654. doi: 10.3390/cells8070654. Cells. 2019. PMID: 31261944 Free PMC article. Review.
References
-
- Abrahamsen, H. N., T. Steiniche, E. Nexo, S. J. Hamilton-Dutoit, and B. S. Sorensen. 2003. Towards quantitative mRNA analysis in paraffin-embedded tissues using real-time reverse transcriptase-polymerase chain reaction: a methodological study on lymph nodes from melanoma patients. J. Mol. Diagn. 5:34-41. - PMC - PubMed
-
- Alamgir, A. S. M., et al. 2000. Phylogenetic analysis of influenza C virus nonstructural (NS) protein genes and identification of the NS2 protein. J. Gen. Virol. 81:1933-1940. - PubMed
-
- Betáková, T., and A. J. Hay. 2007. Evidence that the CM2 protein of influenza C virus can modify the pH of the exocytic pathway of transfected cells. J. Gen. Virol. 88:2291-2296. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
