

Cell type-specific proteasomal processing of HIV-1 Gag-p24 results in an altered epitope repertoire
- PMID: 21106750
- PMCID: PMC3028885
- DOI: 10.1128/JVI.01790-10
Cell type-specific proteasomal processing of HIV-1 Gag-p24 results in an altered epitope repertoire
Abstract
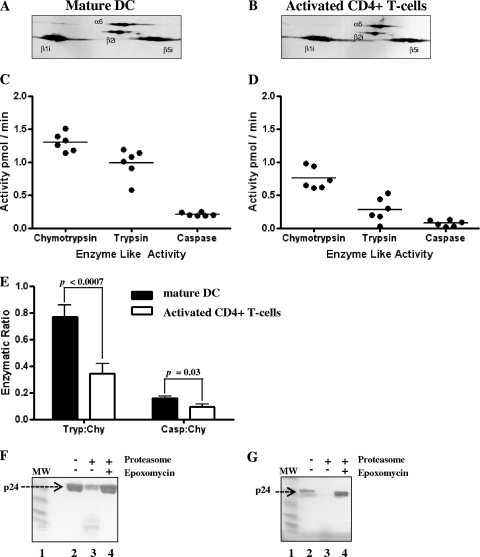
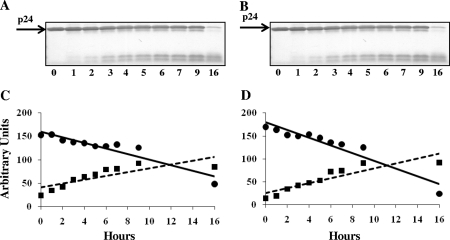
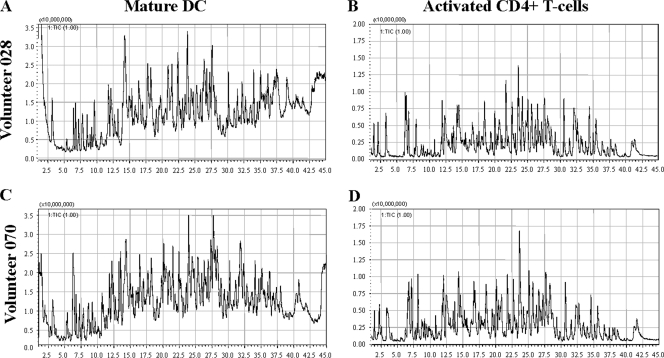
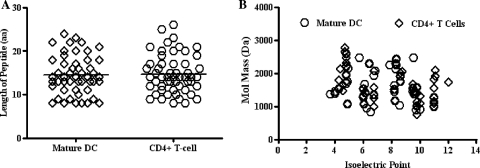
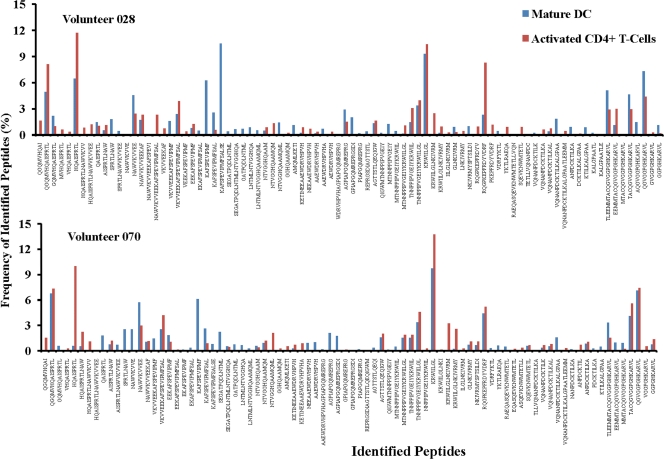
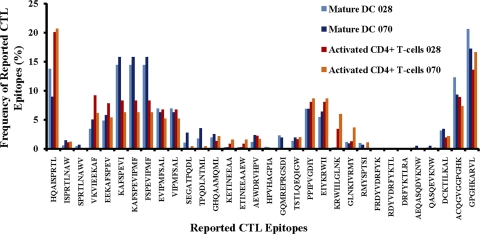
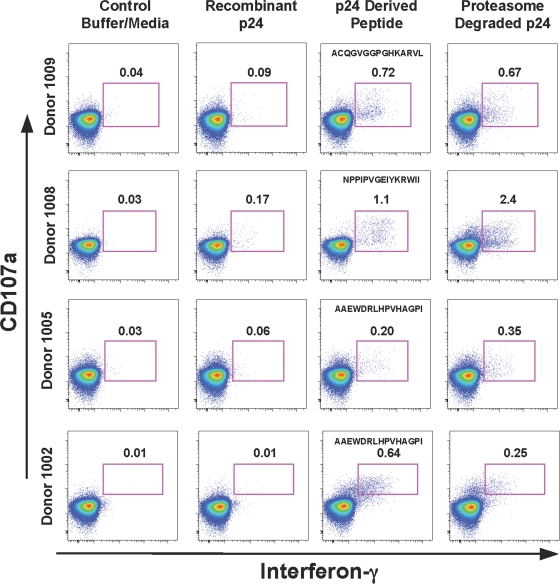
Proteasomes are critical for the processing of antigens for presentation through the major histocompatibility complex (MHC) class I pathway. HIV-1 Gag protein is a component of several experimental HIV-1 vaccines. Therefore, understanding the processing of HIV-1 Gag protein and the resulting epitope repertoire is essential. Purified proteasomes from mature dendritic cells (DC) and activated CD4(+) T cells from the same volunteer were used to cleave full-length Gag-p24 protein, and the resulting peptide fragments were identified by mass spectrometry. Distinct proteasomal degradation patterns and peptide fragments were unique to either mature DC or activated CD4(+) T cells. Almost half of the peptides generated were cell type specific. Two additional differences were observed in the peptides identified from the two cell types. These were in the HLA-B35-Px epitope and the HLA-B27-KK10 epitope. These epitopes have been linked to HIV-1 disease progression. Our results suggest that the source of generation of precursor MHC class I epitopes may be a critical factor for the induction of relevant epitope-specific cytotoxic T cells.
Figures


Similar articles
-
Strategy for identifying dendritic cell-processed CD4+ T cell epitopes from the HIV gag p24 protein.PLoS One. 2012;7(7):e41897. doi: 10.1371/journal.pone.0041897. Epub 2012 Jul 30. PLoS One. 2012. PMID: 22860026 Free PMC article.
-
Heat shock protein-mediated cross-presentation of exogenous HIV antigen on HLA class I and class II.J Immunol. 2004 Aug 1;173(3):1987-93. doi: 10.4049/jimmunol.173.3.1987. J Immunol. 2004. PMID: 15265933
-
HIV-1 variation diminishes CD4 T lymphocyte recognition.J Exp Med. 1998 Nov 16;188(10):1785-93. doi: 10.1084/jem.188.10.1785. J Exp Med. 1998. PMID: 9815256 Free PMC article.
-
Immunodominance of HLA-B27-restricted HIV KK10-specific CD8(+) T-cells is not related to naïve precursor frequency.Immunol Lett. 2013 Jan;149(1-2):119-22. doi: 10.1016/j.imlet.2012.10.002. Epub 2012 Oct 13. Immunol Lett. 2013. PMID: 23068784
-
Cytotoxic T lymphocytes recognize structurally diverse, clade-specific and cross-reactive peptides in human immunodeficiency virus type-1 gag through HLA-B53.Eur J Immunol. 2001 Jun;31(6):1747-56. doi: 10.1002/1521-4141(200106)31:6<1747::aid-immu1747>3.0.co;2-l. Eur J Immunol. 2001. PMID: 11385619
Cited by
-
Analysis of Major Histocompatibility Complex-Bound HIV Peptides Identified from Various Cell Types Reveals Common Nested Peptides and Novel T Cell Responses.J Virol. 2016 Sep 12;90(19):8605-20. doi: 10.1128/JVI.00599-16. Print 2016 Oct 1. J Virol. 2016. PMID: 27440904 Free PMC article.
-
Latency reversal agents modulate HIV antigen processing and presentation to CD8 T cells.PLoS Pathog. 2020 Mar 20;16(3):e1008442. doi: 10.1371/journal.ppat.1008442. eCollection 2020 Mar. PLoS Pathog. 2020. PMID: 32196533 Free PMC article.
-
Antigen processing and presentation in HIV infection.Mol Immunol. 2019 Sep;113:67-74. doi: 10.1016/j.molimm.2018.03.027. Epub 2018 Apr 7. Mol Immunol. 2019. PMID: 29636181 Free PMC article. Review.
-
Rapid antigen processing and presentation of a protective and immunodominant HLA-B*27-restricted hepatitis C virus-specific CD8+ T-cell epitope.PLoS Pathog. 2012;8(11):e1003042. doi: 10.1371/journal.ppat.1003042. Epub 2012 Nov 29. PLoS Pathog. 2012. PMID: 23209413 Free PMC article. Clinical Trial.
-
Computational and experimental validation of B and T-cell epitopes of the in vivo immune response to a novel malarial antigen.PLoS One. 2013 Aug 16;8(8):e71610. doi: 10.1371/journal.pone.0071610. eCollection 2013. PLoS One. 2013. PMID: 23977087 Free PMC article.
References
-
- Ackerman, A. L., A. Giodini, and P. Cresswell. 2006. A role for the endoplasmic reticulum protein retrotranslocation machinery during crosspresentation by dendritic cells. Immunity 25:607-617. - PubMed
-
- Altfeld, M., et al. 2003. Influence of HLA-B57 on clinical presentation and viral control during acute HIV-1 infection. AIDS 17:2581-2591. - PubMed
-
- Basler, M., N. Youhnovski, M. Van Den Broek, M. Przybylski, and M. Groettrup. 2004. Immunoproteasomes down-regulate presentation of a subdominant T cell epitope from lymphocytic choriomeningitis virus. J. Immunol. 173:3925-3934. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
