

The absence of eye muscle fatigue indicates that the nervous system compensates for non-motor disturbances of oculomotor function
- PMID: 21106822
- PMCID: PMC6633742
- DOI: 10.1523/JNEUROSCI.3901-10.2010
The absence of eye muscle fatigue indicates that the nervous system compensates for non-motor disturbances of oculomotor function
Abstract
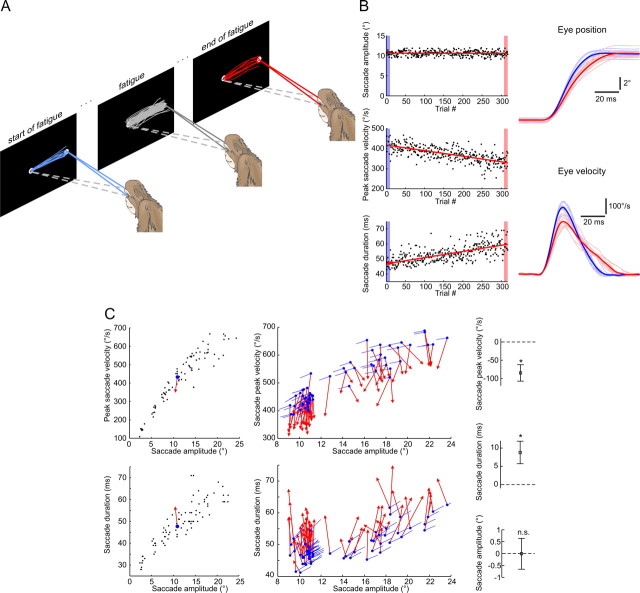
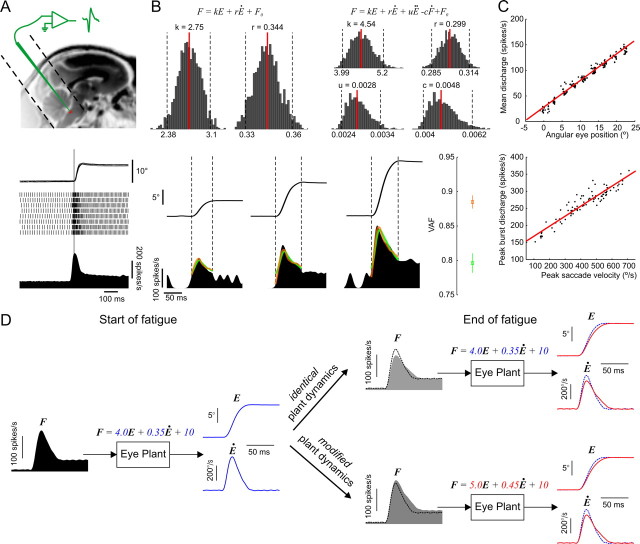
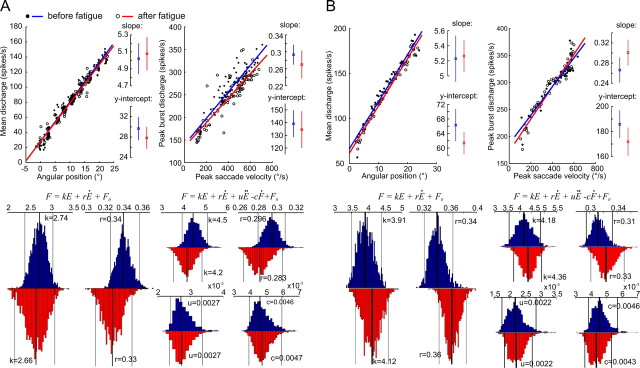
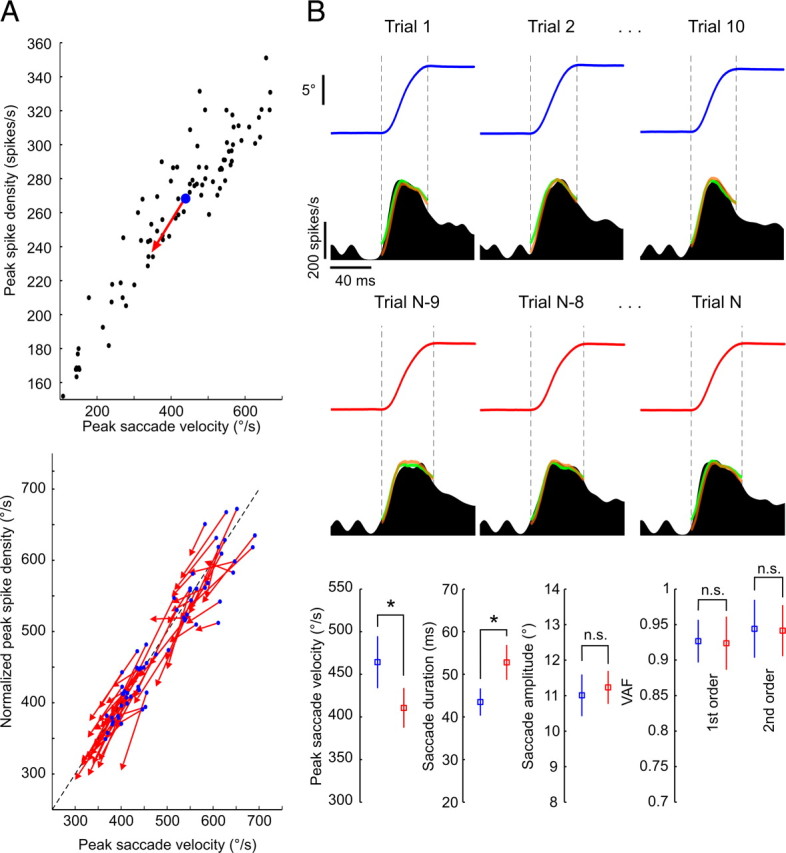
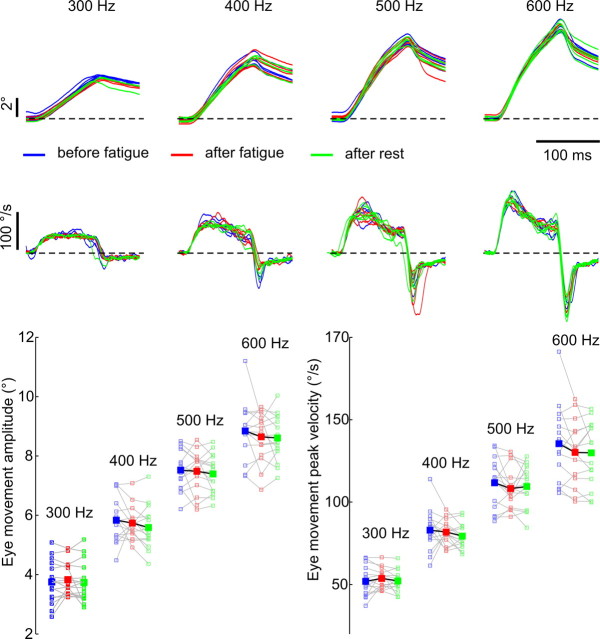
The physical properties of our bodies are subject to change (due to fatigue, heavy equipment, injury or aging) as we move around in the surrounding environment. The traditional definition of motor adaptation dictates that a mechanism in our brain needs to compensate for such alterations by appropriately modifying neural motor commands, if the vitally important accuracy of executed movements is to be preserved. In this article we describe how a repetitive eye movement task brings about changes in eye saccade kinematics that compromise accurate motor performance in the absence of a proper compensatory response. Surgical lesions in animals and human patient studies have previously demonstrated that an intact cerebellum is necessary for the compensation to arise and prevent the occurrence of hypometric movements. Here we identified the dynamic properties of the eye plant by recording from abducens motoneurons responsible for the required movement and measured the muscle response to microstimulation of the abducens nucleus in rhesus monkeys. The ensuing results demonstrate that the muscular periphery remains intact during the fatiguing eye movement task, while internal sources of noise (drowsiness, attentional modulation, neuronal fatigue etc.) must be responsible for a diminished oculomotor performance. This finding leads to the important realization that while supervising the accuracy of our movements, the nervous system takes additionally into account and adapts to any disruptive processes within the brain itself, clearly unrelated to the dynamical behavior of muscles or the environment. The existence of this supplementary mechanism forces a reassessment of traditional views of cerebellum-dependent motor adaptation.
Figures
References
-
- Abel LA, Schmidt D, Dell'Osso LF, Daroff RB. Saccadic system plasticity in humans. Ann Neurol. 1978;4:313–318. - PubMed
-
- Bahill AT, Stark L. Overlapping saccades and glissades are produced by fatigue in saccadic eye-movement system. Exp Neurol. 1975;48:95–106. - PubMed
-
- Brozek J. Quantitative criteria of oculomotor performance and fatigue. J Appl Physiol. 1949;2:247–260. - PubMed
-
- Büttner-Ennever JA, Horn AK, Scherberger H, D'Ascanio P. Motoneurons of twitch and nontwitch extraocular muscle fibers in the abducens, trochlear, and oculomotor nuclei of monkeys. J Comp Neurol. 2001;438:318–335. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources