

A mitochondrial enzyme degrades carotenoids and protects against oxidative stress
- PMID: 21106934
- PMCID: PMC3042841
- DOI: 10.1096/fj.10-173906
A mitochondrial enzyme degrades carotenoids and protects against oxidative stress
Abstract
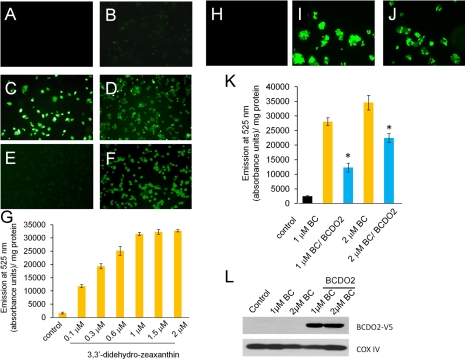
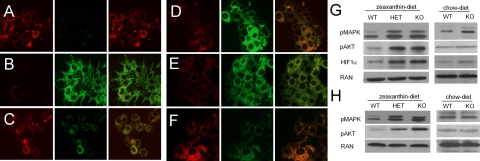
Carotenoids are the precursors for vitamin A and are proposed to prevent oxidative damage to cells. Mammalian genomes encode a family of structurally related nonheme iron oxygenases that modify double bonds of these compounds by oxidative cleavage and cis-to-trans isomerization. The roles of the family members BCMO1 and RPE65 for vitamin A production and vision have been well established. Surprisingly, we found that the third family member, β,β-carotene-9',10'-oxygenase (BCDO2), is a mitochondrial carotenoid-oxygenase with broad substrate specificity. In BCDO2-deficient mice, carotenoid homeostasis was abrogated, and carotenoids accumulated in several tissues. In hepatic mitochondria, accumulated carotenoids induced key markers of mitochondrial dysfunction, such as manganese superoxide dismutase (9-fold), and reduced rates of ADP-dependent respiration by 30%. This impairment was associated with an 8- to 9-fold induction of phosphor-MAP kinase and phosphor-AKT, markers of cell signaling pathways related to oxidative stress and disease. Administration of carotenoids to human HepG2 cells depolarized mitochondrial membranes and resulted in the production of reactive oxygen species. Thus, our studies in BCDO2-deficient mice and human cell cultures indicate that carotenoids can impair respiration and induce oxidative stress. Mammalian cells thus express a mitochondrial carotenoid-oxygenase that degrades carotenoids to protect these vital organelles.
Figures
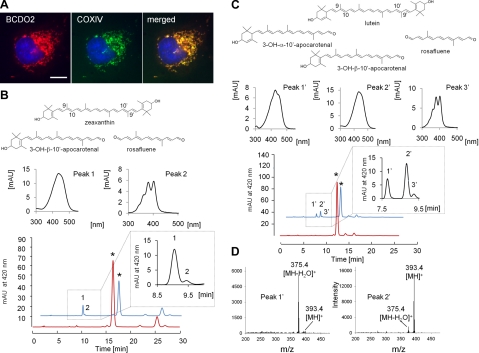
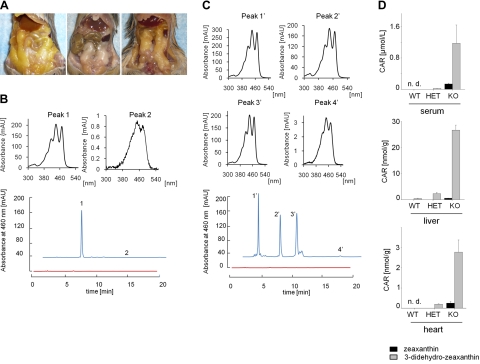
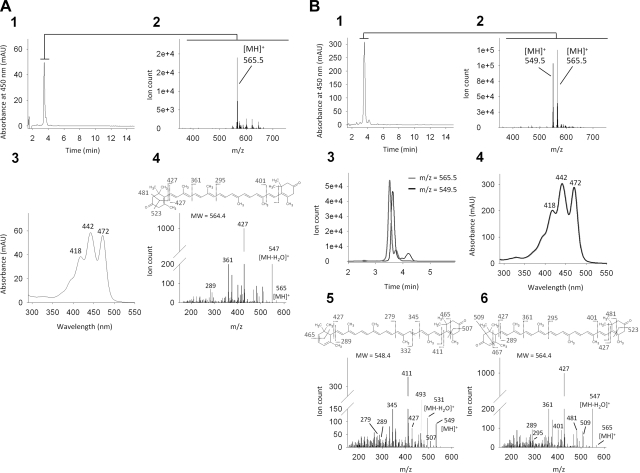
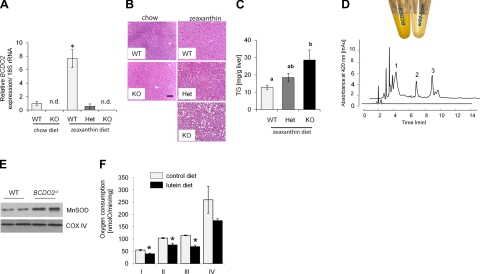
References
-
- Demmig-Adams B., Adams W. W., 3rd. (2002) Antioxidants in photosynthesis and human nutrition. Science 298, 2149–2153 - PubMed
-
- Bernstein P. S., Yoshida M. D., Katz N. B., McClane R. W., Gellermann W. (1998) Raman detection of macular carotenoid pigments in intact human retina. Invest. Ophthalmol. Vis. Sci. 39, 2003–2011 - PubMed
-
- Snodderly D. M., Handelman G. J., Adler A. J. (1991) Distribution of individual macular pigment carotenoids in central retina of macaque and squirrel monkeys. Invest. Ophthalmol. Vis. Sci. 32, 268–279 - PubMed
-
- Krinsky N. I., Landrum J. T., Bone R. A. (2003) Biologic mechanisms of the protective role of lutein and zeaxanthin in the eye. Annu. Rev. Nutr. 23, 171–201 - PubMed
-
- Moise A. R., von Lintig J., Palczewski K. (2005) Related enzymes solve evolutionarily recurrent problems in the metabolism of carotenoids. Trends Plant Sci. 10, 178–186 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
