

Implications of the polymorphism of HLA-G on its function, regulation, evolution and disease association
- PMID: 21107637
- PMCID: PMC3021195
- DOI: 10.1007/s00018-010-0580-7
Implications of the polymorphism of HLA-G on its function, regulation, evolution and disease association
Abstract
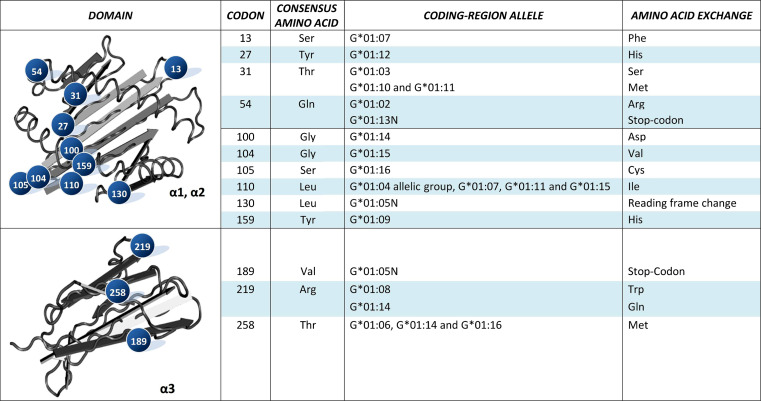
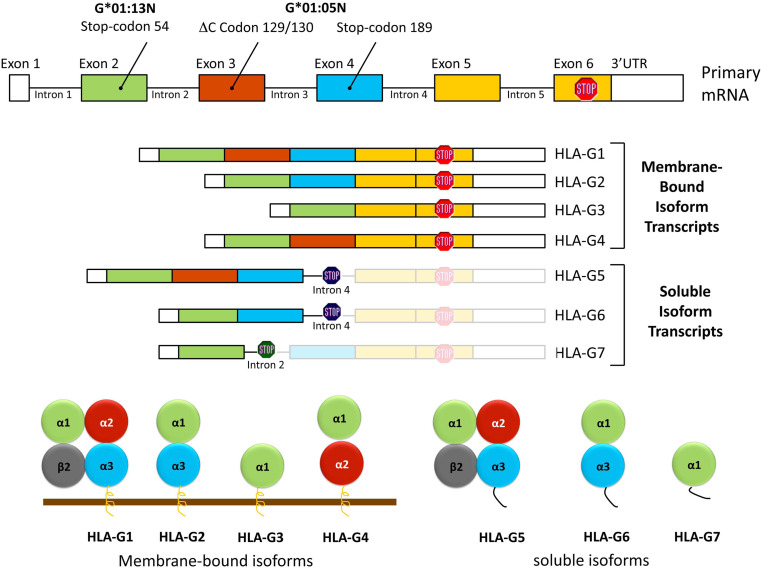
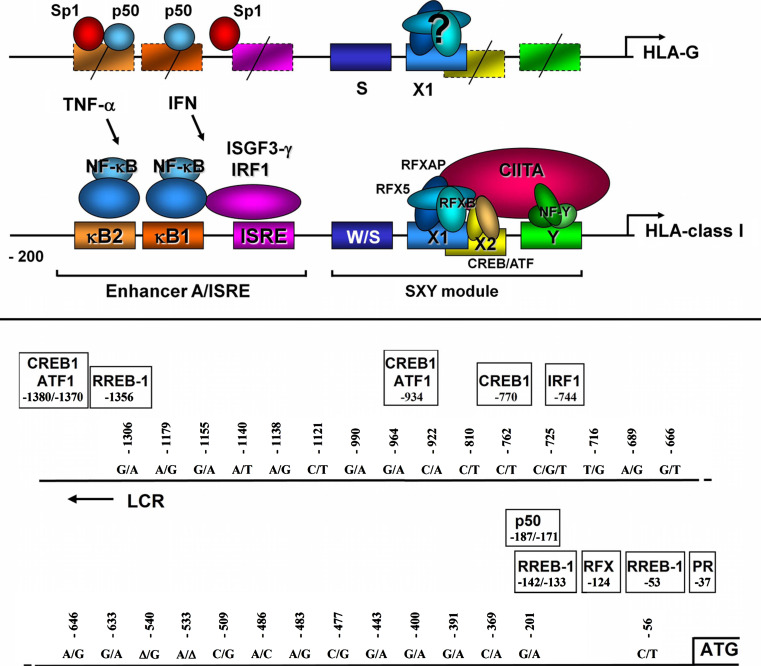
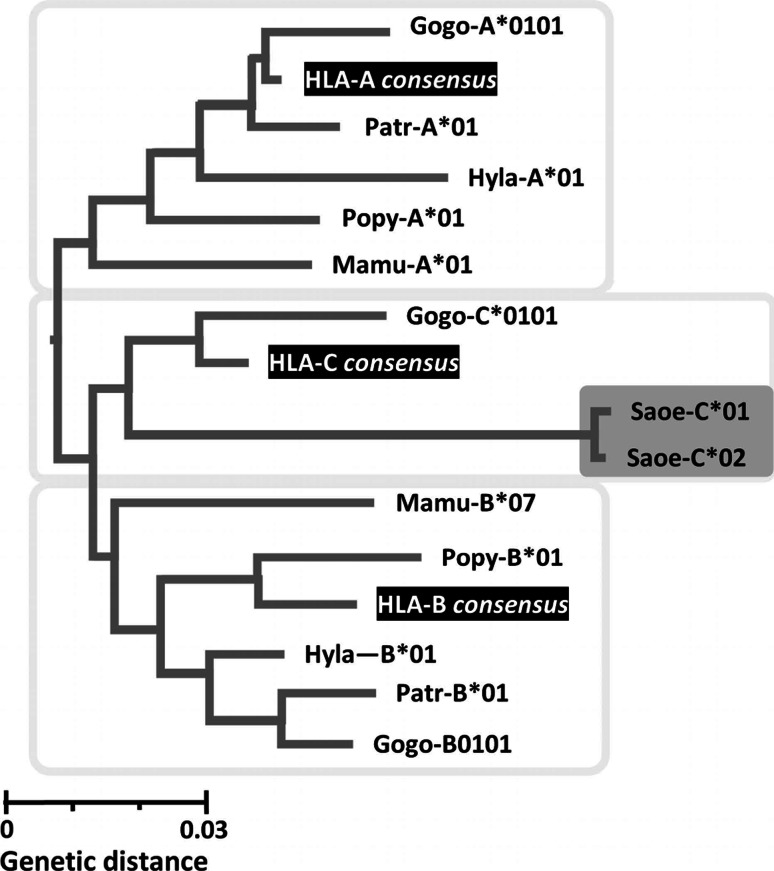
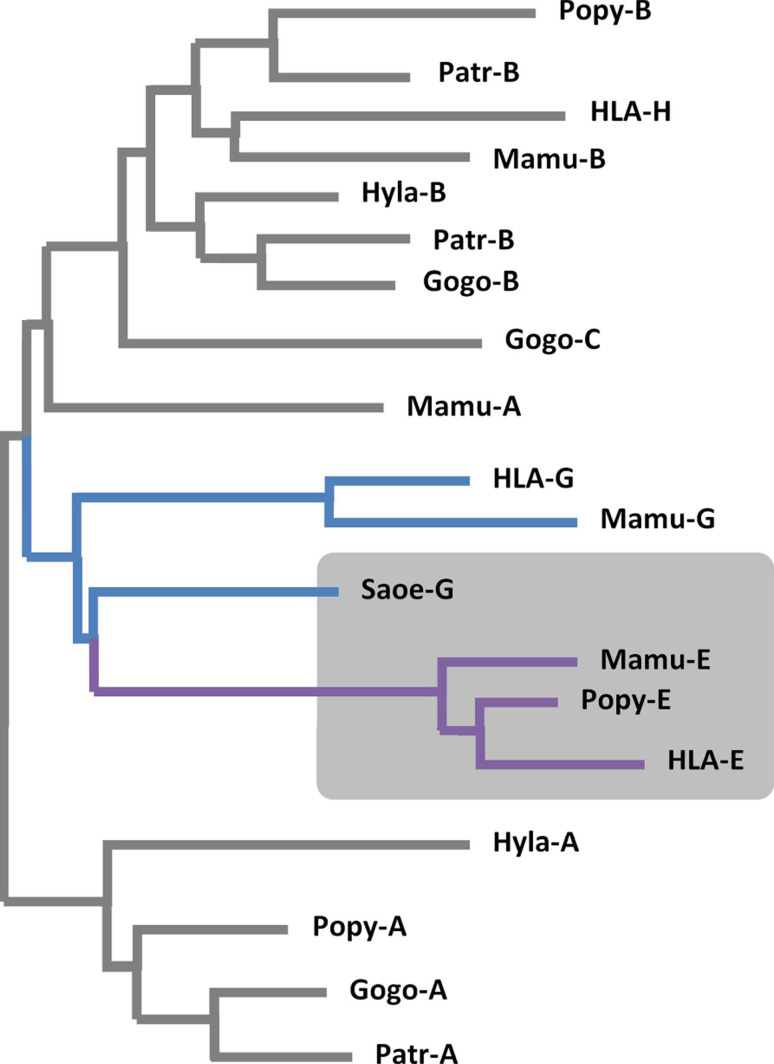
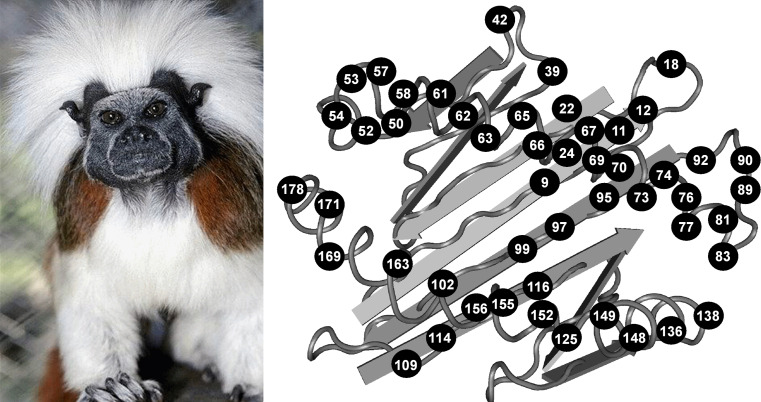
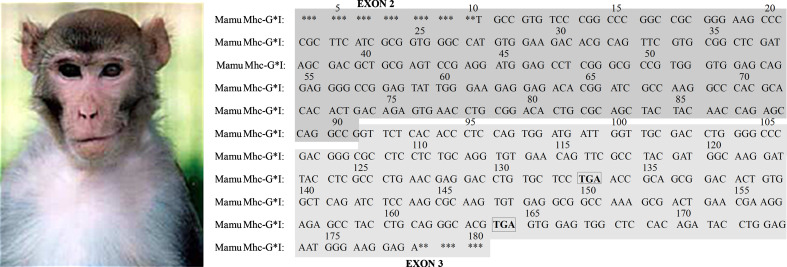
The HLA-G gene displays several peculiarities that are distinct from those of classical HLA class I genes. The unique structure of the HLA-G molecule permits a restricted peptide presentation and allows the modulation of the cells of the immune system. Although polymorphic sites may potentially influence all biological functions of HLA-G, those present at the promoter and 3' untranslated regions have been particularly studied in experimental and pathological conditions. The relatively low polymorphism observed in the MHC-G coding region both in humans and apes may represent a strong selective pressure for invariance, whereas, in regulatory regions several lines of evidence support the role of balancing selection. Since HLA-G has immunomodulatory properties, the understanding of gene regulation and the role of polymorphic sites on gene function may permit an individualized approach for the future use of HLA-G for therapeutic purposes.
Figures




References
-
- Ishitani A, Sageshima N, Lee N, Dorofeeva N, Hatake K, Marquardt H, Geraghty DE. Protein expression and peptide binding suggest unique and interacting functional roles for HLA-E, F, and G in maternal-placental immune recognition. J Immunol. 2003;171:1376–1384. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
