

DPI-ELISA: a fast and versatile method to specify the binding of plant transcription factors to DNA in vitro
- PMID: 21108821
- PMCID: PMC3003642
- DOI: 10.1186/1746-4811-6-25
DPI-ELISA: a fast and versatile method to specify the binding of plant transcription factors to DNA in vitro
Abstract
Background: About 10% of all genes in eukaryote genomes are predicted to encode transcription factors. The specific binding of transcription factors to short DNA-motifs influences the expression of neighbouring genes. However, little is known about the DNA-protein interaction itself. To date there are only a few suitable methods to characterise DNA-protein-interactions, among which the EMSA is the method most frequently used in laboratories. Besides EMSA, several protocols describe the effective use of an ELISA-based transcription factor binding assay e.g. for the analysis of human NFκB binding to specific DNA sequences.
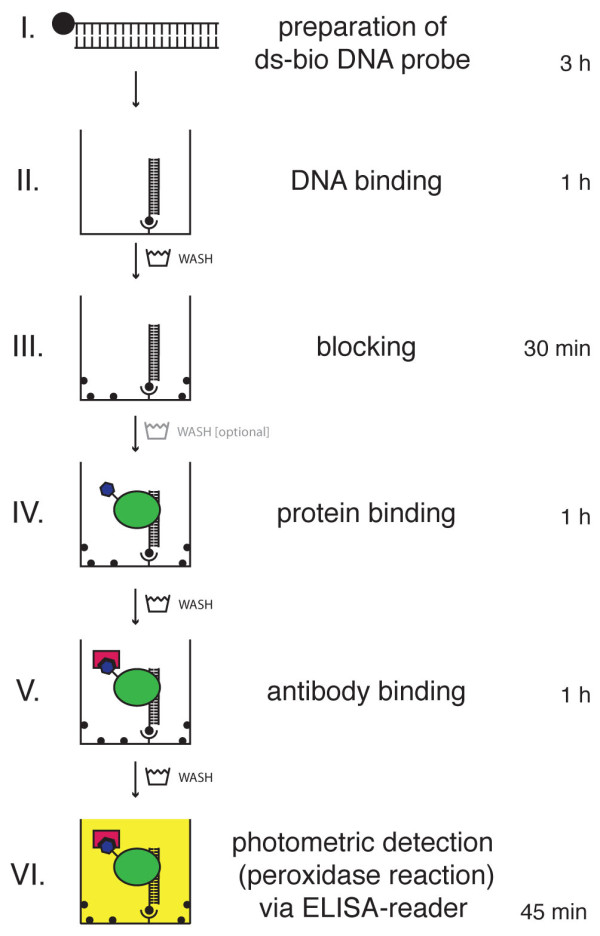
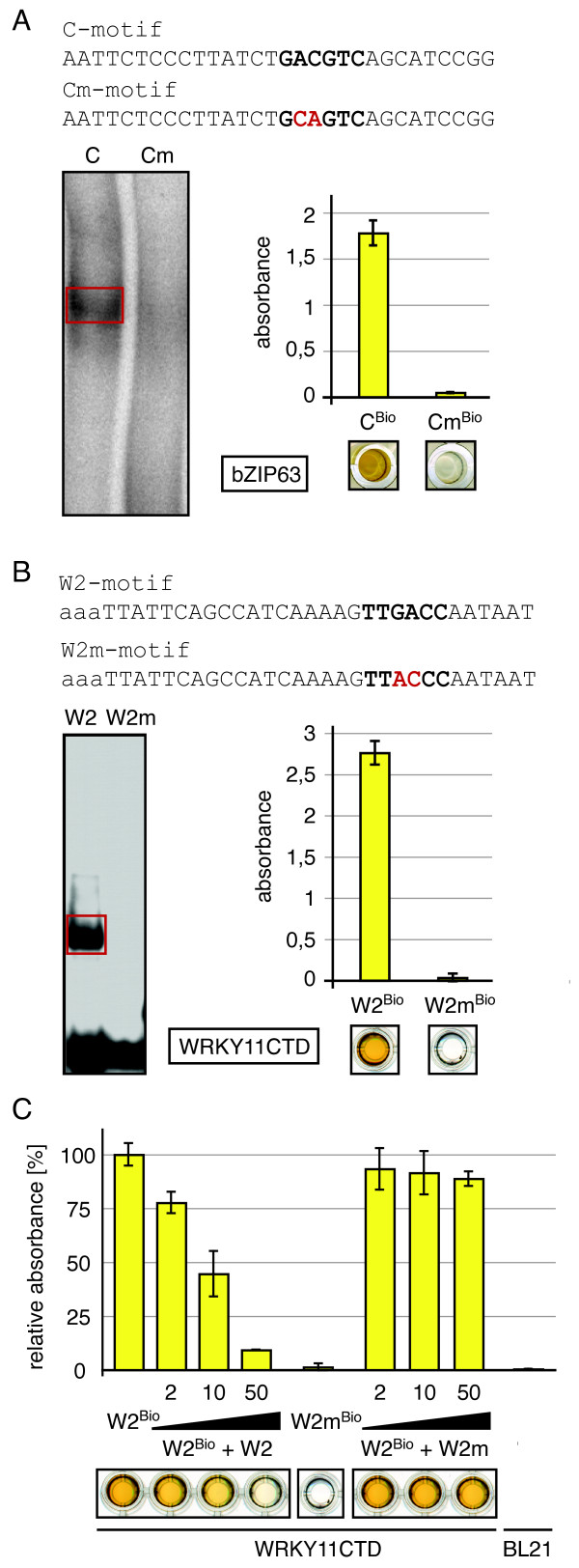
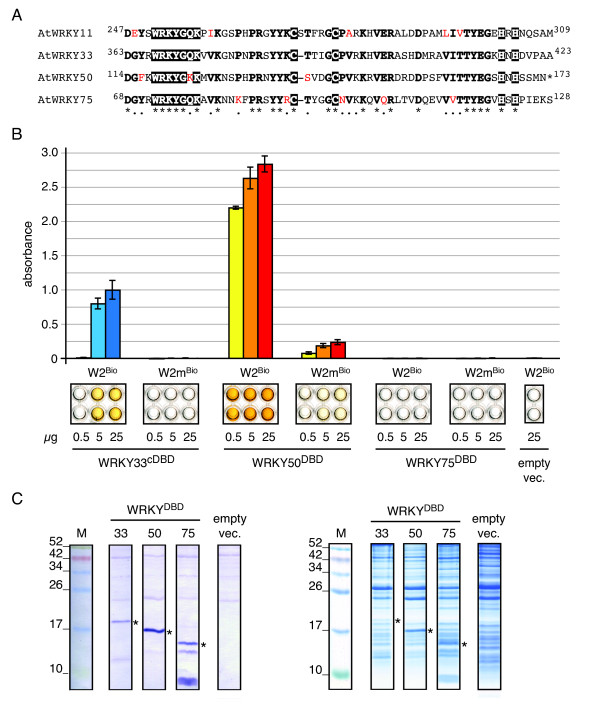
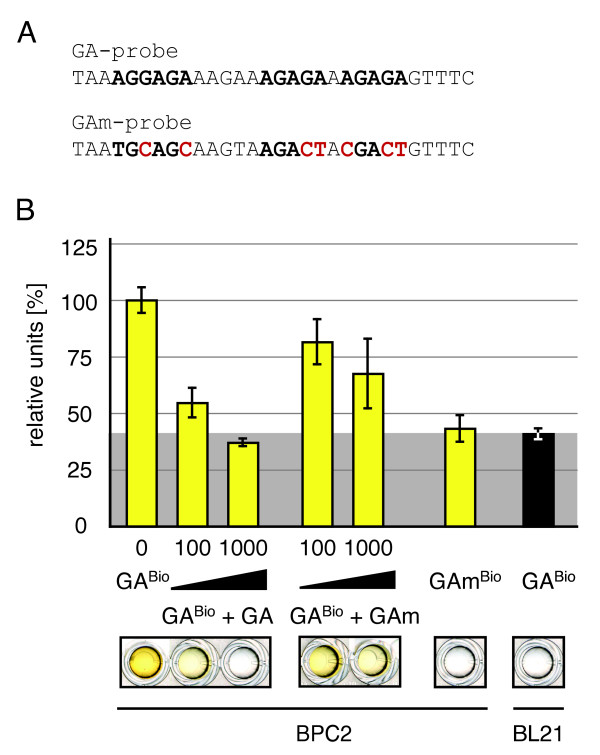
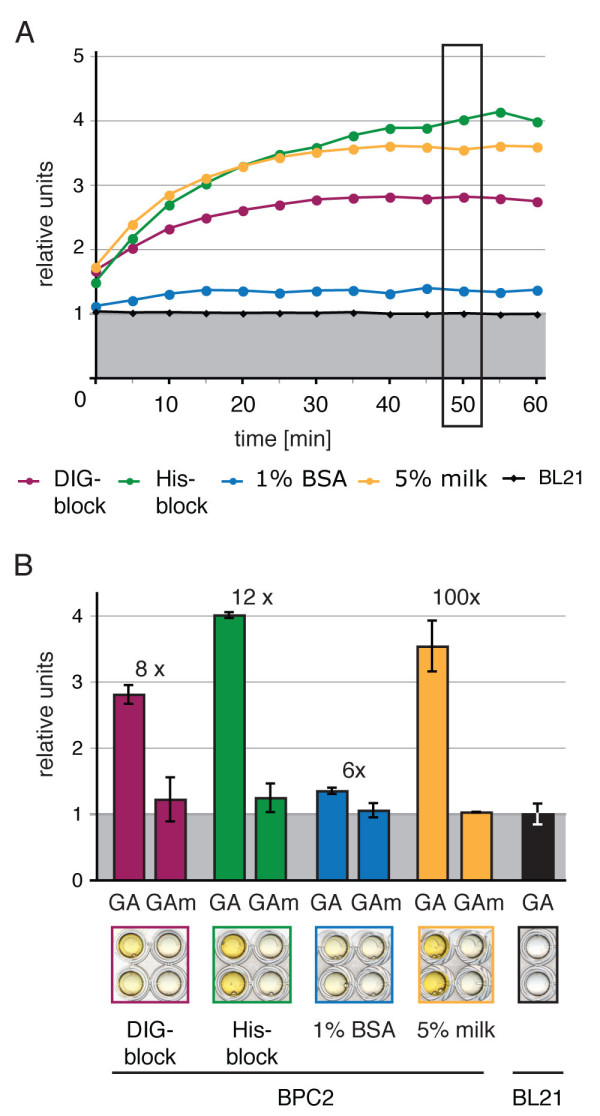
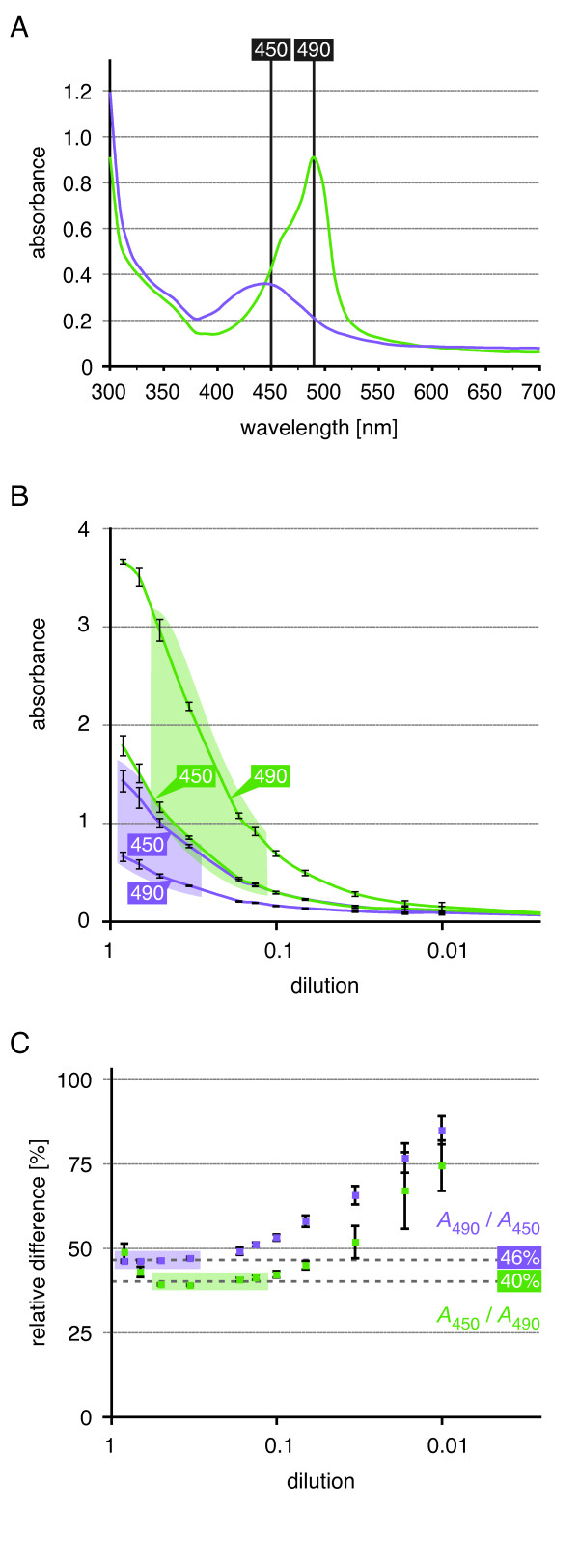
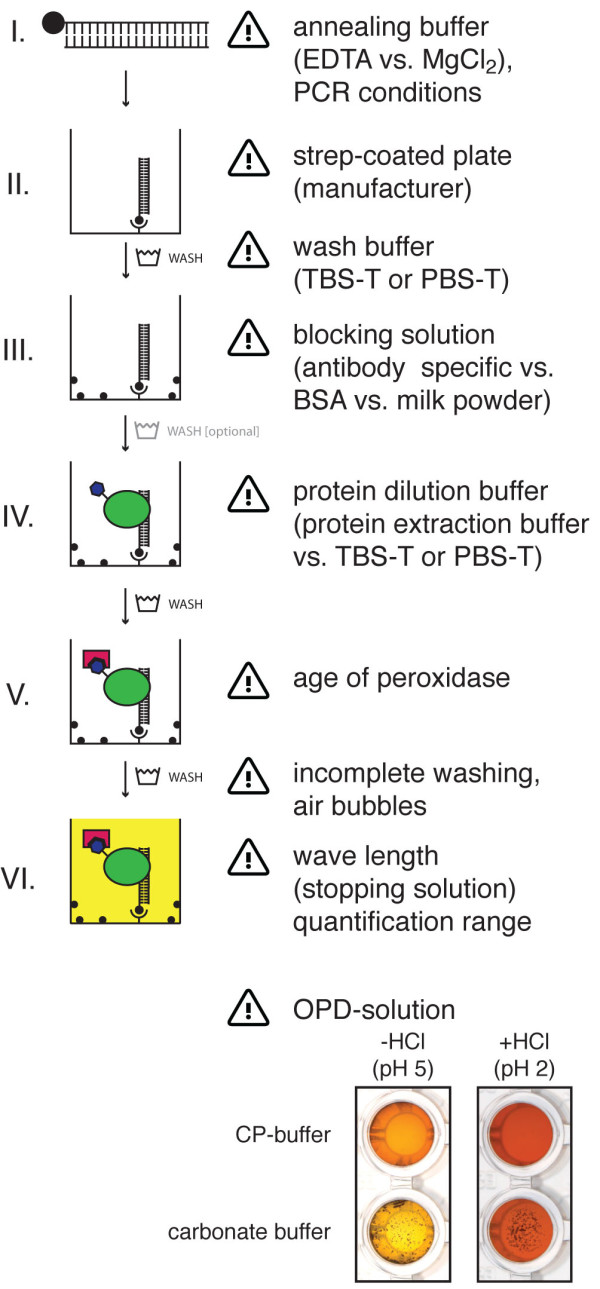
Results: We provide a unified protocol for this type of ELISA analysis, termed DNA-Protein-Interaction (DPI)-ELISA. Qualitative analyses with His-epitope tagged plant transcription factors expressed in E. coli revealed that EMSA and DPI-ELISA result in comparable and reproducible data. The binding of AtbZIP63 to the C-box and AtWRKY11 to the W2-box could be reproduced and validated by both methods. We next examined the physical binding of the C-terminal DNA-binding domains of AtWRKY33, AtWRKY50 and AtWRKY75 to the W2-box. Although the DNA-binding domain is highly conserved among the WRKY proteins tested, the use of the DPI-ELISA discloses differences in W2-box binding properties between these proteins. In addition to these well-studied transcription factor families, we applied our protocol to AtBPC2, a member of the so far uncharacterised plant specific Basic Pentacysteine transcription factor family. We could demonstrate binding to GA/TC-dinucleotide repeat motifs by our DPI-ELISA protocol. Different buffers and reaction conditions were examined.
Conclusions: We successfully applied our DPI-ELISA protocol to investigate the DNA-binding specificities of three different classes of transcription factors from Arabidopsis thaliana. However, the analysis of the binding affinity of any DNA-binding protein to any given DNA sequence can be performed via this method. The DPI-ELISA is cost efficient, less time-consuming than other methods and provides a qualitative and quantitative readout. The presented DPI-ELISA protocol is accompanied by advice on trouble-shooting, which will enable scientists to rapidly establish this versatile and easy to use method in their laboratories.
Figures
References
-
- Berendzen KW, Harter K, Wanke D. Analysis of plant regulatory DNA sequences by transient protoplast assays and computer aided sequence evaluation. Methods Mol Biol. 2009;479:311–335. - PubMed
-
- Wanke D, Harter K. Analysis of plant regulatory DNA sequences by the yeast-one-hybrid assay. Methods Mol Biol. 2009;479:291–309. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
