

Function and evolution of sex determination mechanisms, genes and pathways in insects
- PMID: 21110346
- PMCID: PMC3040292
- DOI: 10.1002/bies.201000043
Function and evolution of sex determination mechanisms, genes and pathways in insects
Abstract
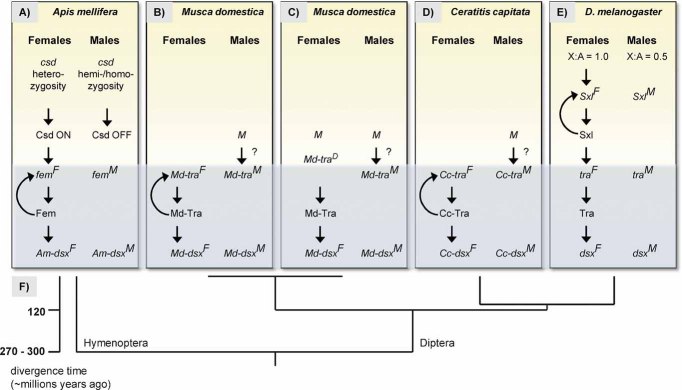
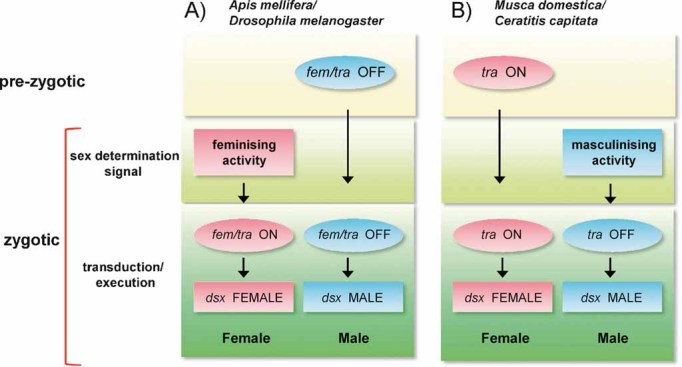
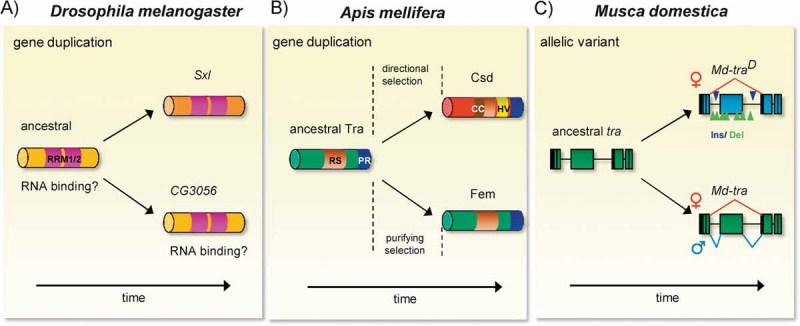
Animals have evolved a bewildering diversity of mechanisms to determine the two sexes. Studies of sex determination genes--their history and function--in non-model insects and Drosophila have allowed us to begin to understand the generation of sex determination diversity. One common theme from these studies is that evolved mechanisms produce activities in either males or females to control a shared gene switch that regulates sexual development. Only a few small-scale changes in existing and duplicated genes are sufficient to generate large differences in sex determination systems. This review summarises recent findings in insects, surveys evidence of how and why sex determination mechanisms can change rapidly and suggests fruitful areas of future research.
Figures
References
-
- Cline TW, Meyer BJ. Vive la difference: males vs. females in flies vs. worms. Annu Rev Genet. 1996;30:637–702. - PubMed
-
- Lovell-Badge R, Canning C, Sekido R. Sex-determining genes in mice: building pathways. Novartis Found Symp. 2002;244:4–18. - PubMed
-
- Sekido R, Lovell-Badge R. Sex determination involves synergistic action of SRY and SF1 on a specific Sox9 enhancer. Nature. 2008;453:930–4. - PubMed
-
- Marshall Graves JA. Weird animal genomes and the evolution of vertebrate sex and sex chromosomes. Annu Rev Genet. 2008;42:565–86. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
