

Abortive HIV infection mediates CD4 T cell depletion and inflammation in human lymphoid tissue
- PMID: 21111238
- PMCID: PMC3026834
- DOI: 10.1016/j.cell.2010.11.001
Abortive HIV infection mediates CD4 T cell depletion and inflammation in human lymphoid tissue
Erratum in
- Cell. 2014 Feb 27;156(5):1112-3
Abstract
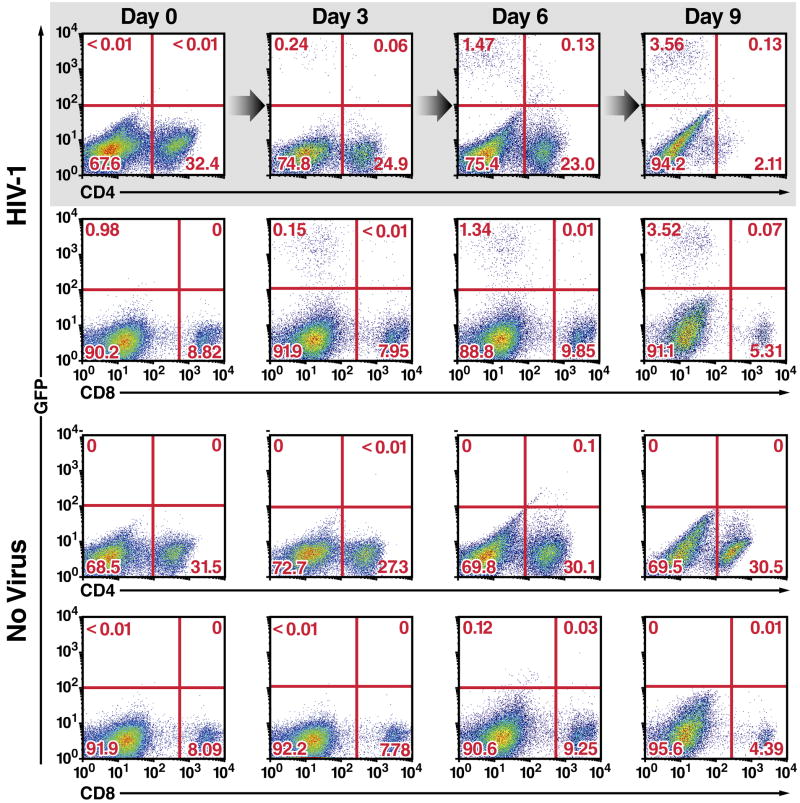
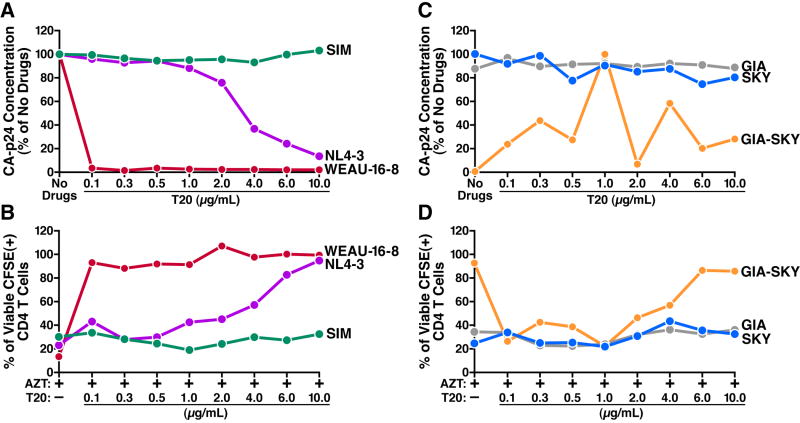
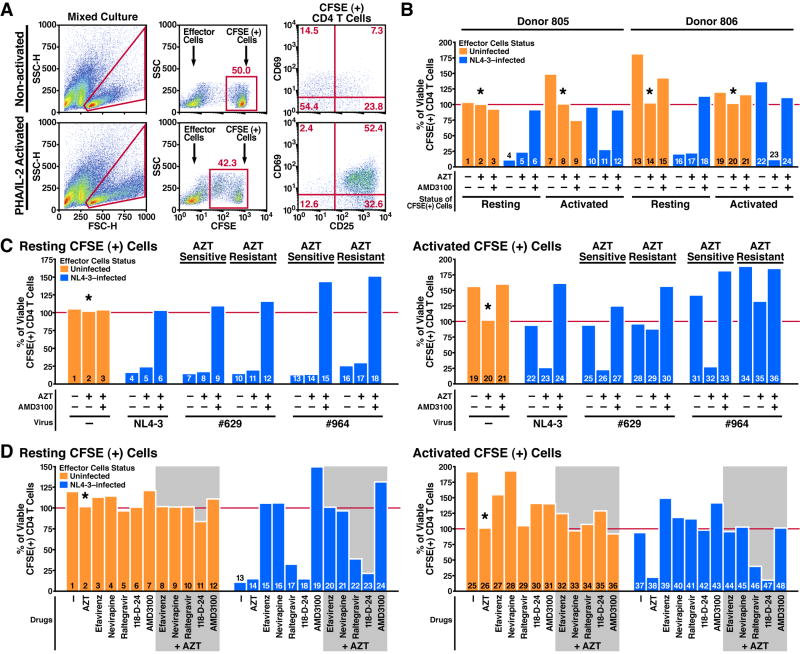
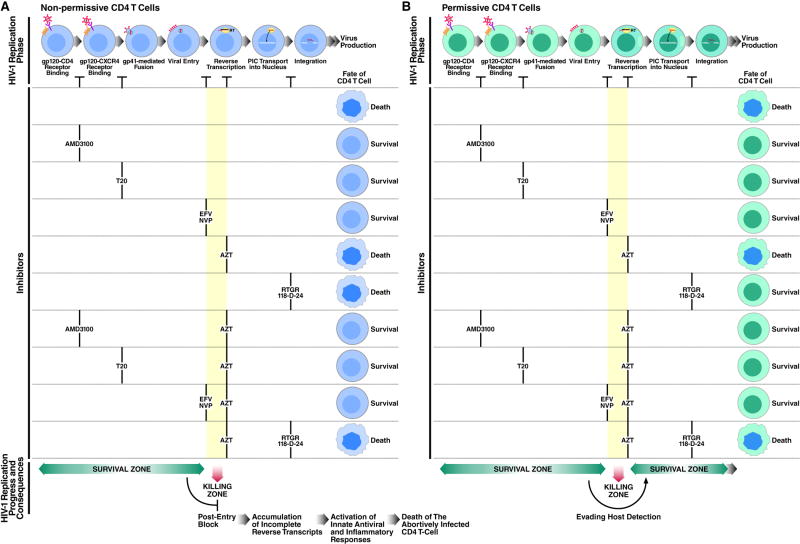
The mechanism by which CD4 T cells are depleted in HIV-infected hosts remains poorly understood. In ex vivo cultures of human tonsil tissue, CD4 T cells undergo a pronounced cytopathic response following HIV infection. Strikingly, >95% of these dying cells are not productively infected but instead correspond to bystander cells. We now show that the death of these "bystander" cells involves abortive HIV infection. Inhibitors blocking HIV entry or early steps of reverse transcription prevent CD4 T cell death while inhibition of later events in the viral life cycle does not. We demonstrate that the nonpermissive state exhibited by the majority of resting CD4 tonsil T cells leads to accumulation of incomplete reverse transcripts. These cytoplasmic nucleic acids activate a host defense program that elicits a coordinated proapoptotic and proinflammatory response involving caspase-3 and caspase-1 activation. While this response likely evolved to protect the host, it centrally contributes to the immunopathogenic effects of HIV.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures



References
-
- Cavrois M, De Noronha C, Greene WC. A sensitive and specific enzyme-based assay detecting HIV-1 virion fusion in primary T lymphocytes. Nat Biotechnol. 2002;20:1151–1154. - PubMed
-
- Eckstein DA, Penn ML, Korin YD, Scripture-Adams DD, Zack JA, Kreisberg JF, Roederer M, Sherman MP, Chin PS, Goldsmith MA. HIV-1 actively replicates in naive CD4(+) T cells residing within human lymphoid tissues. Immunity. 2001;15:671–682. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
