

FOXO/4E-BP signaling in Drosophila muscles regulates organism-wide proteostasis during aging
- PMID: 21111239
- PMCID: PMC3066043
- DOI: 10.1016/j.cell.2010.10.007
FOXO/4E-BP signaling in Drosophila muscles regulates organism-wide proteostasis during aging
Abstract
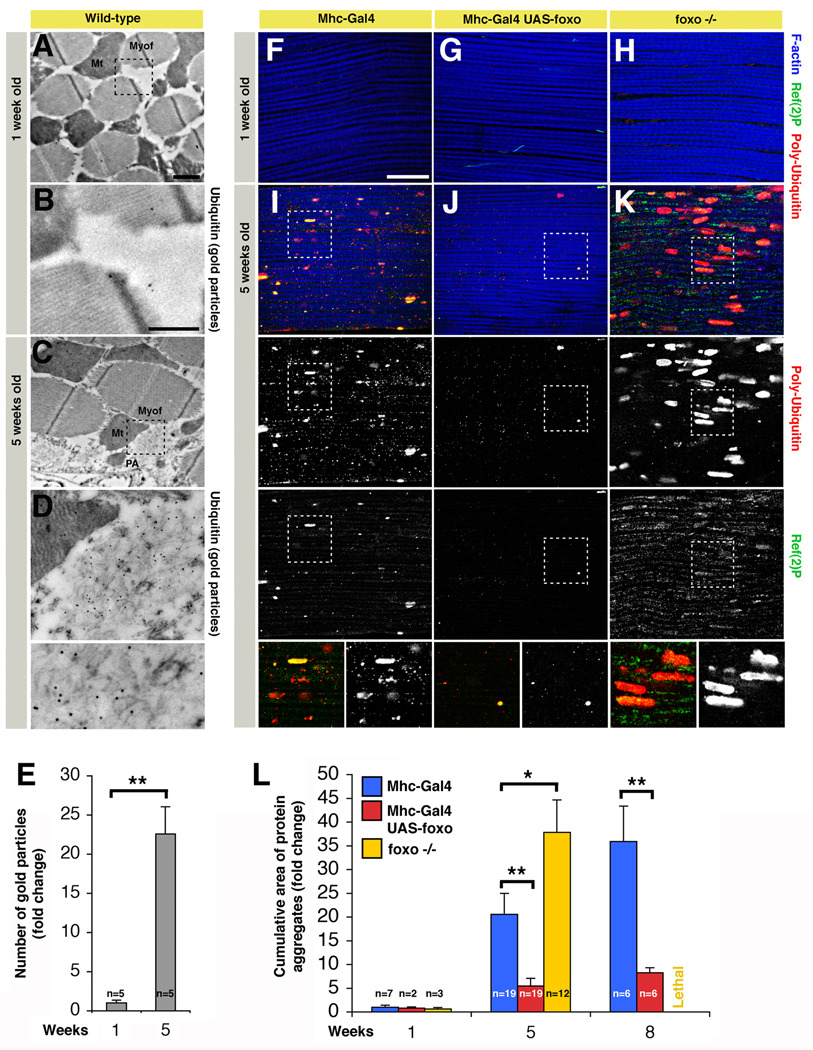
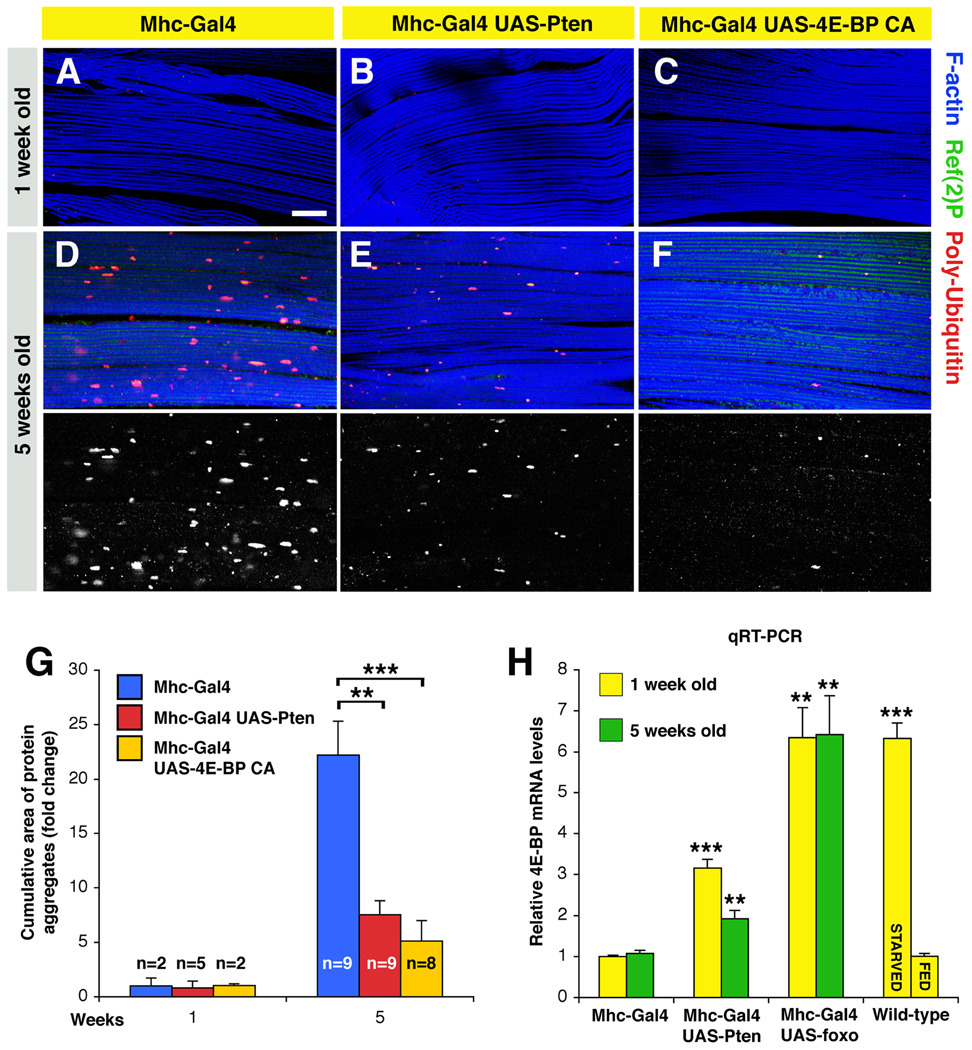
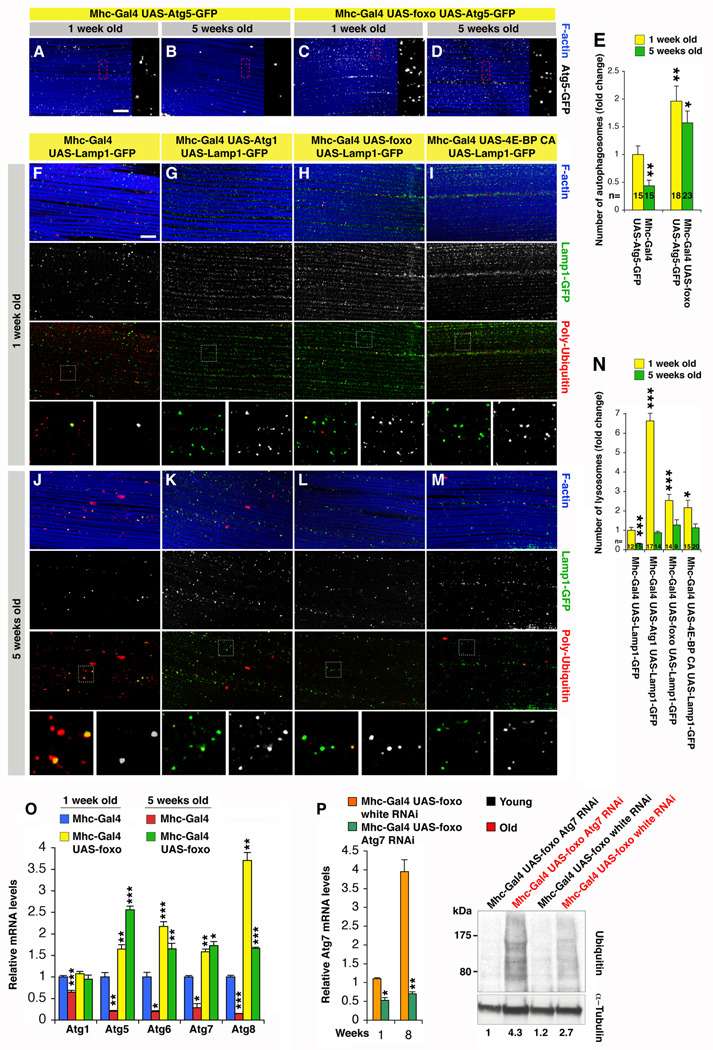
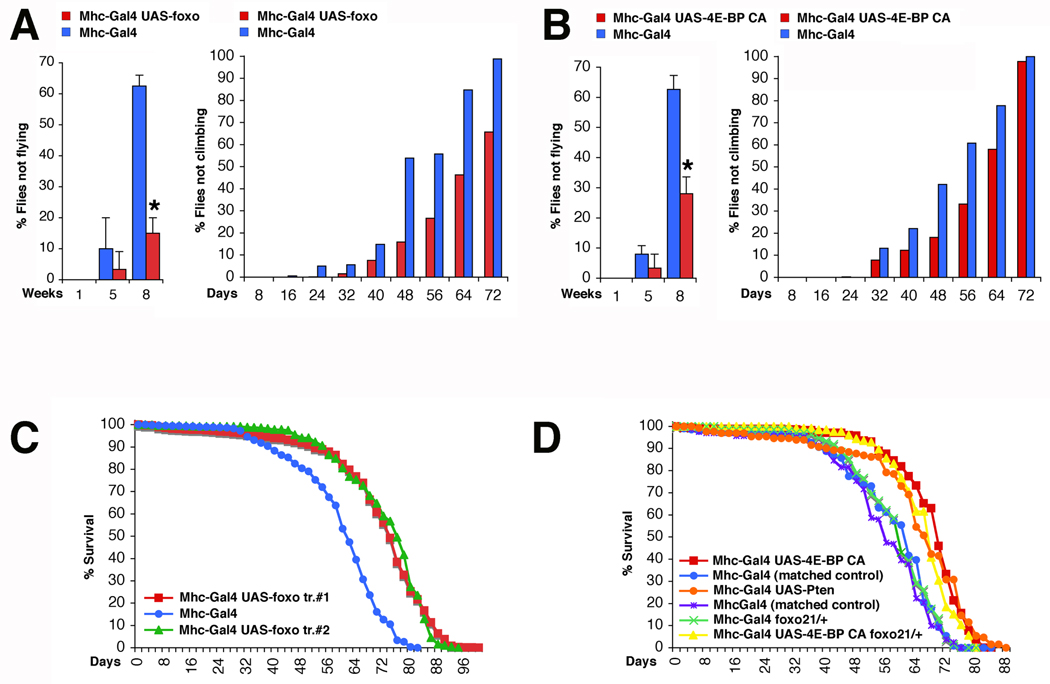
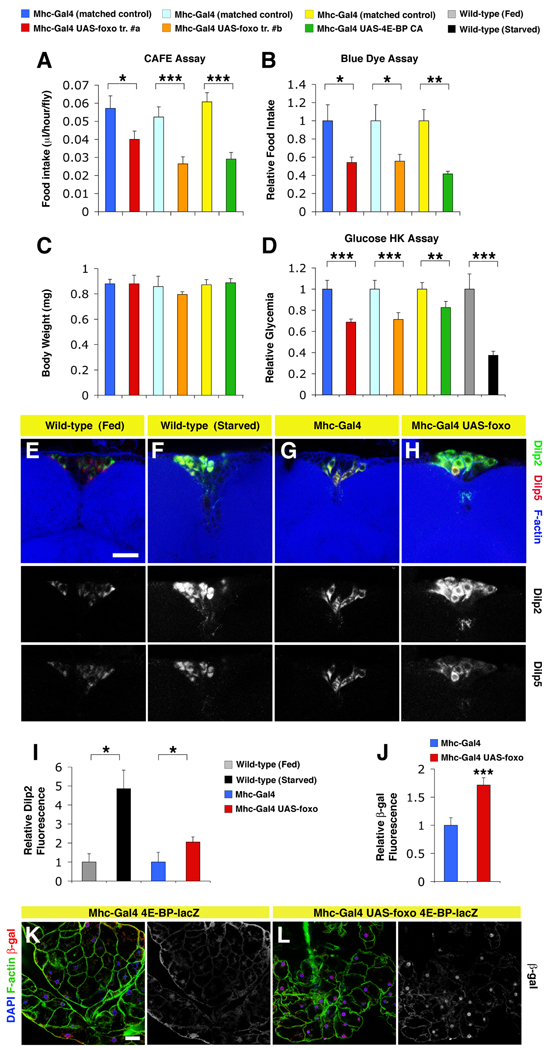
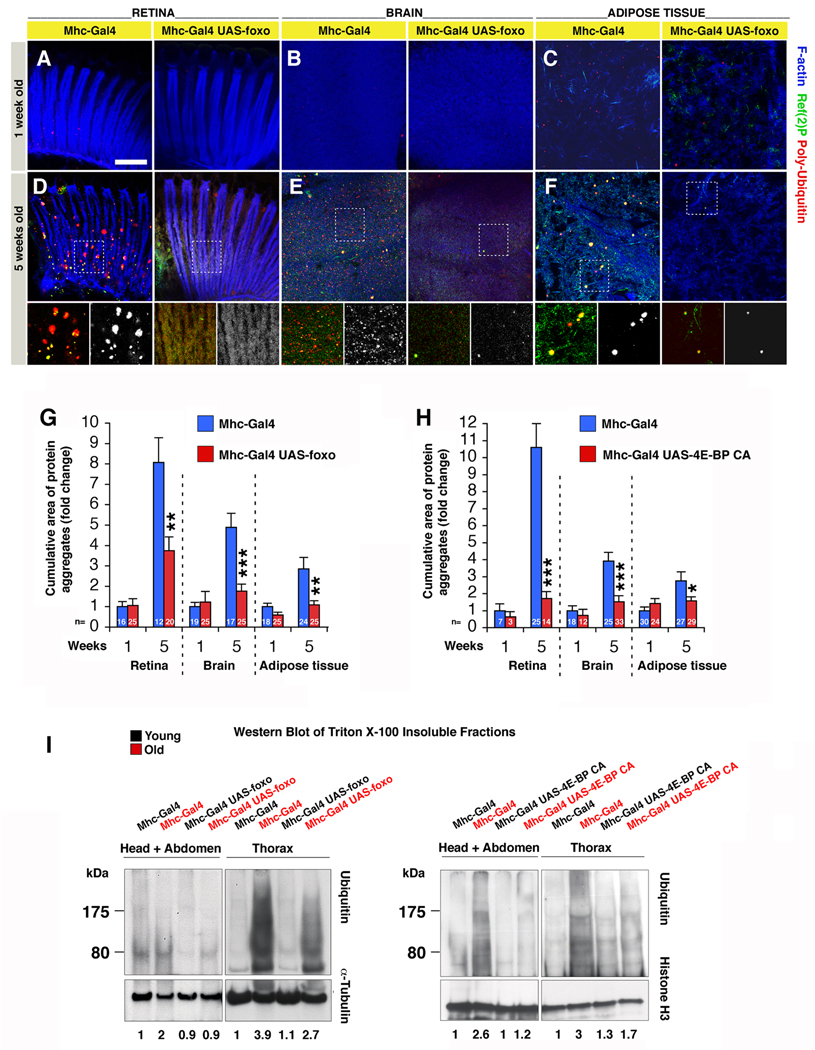
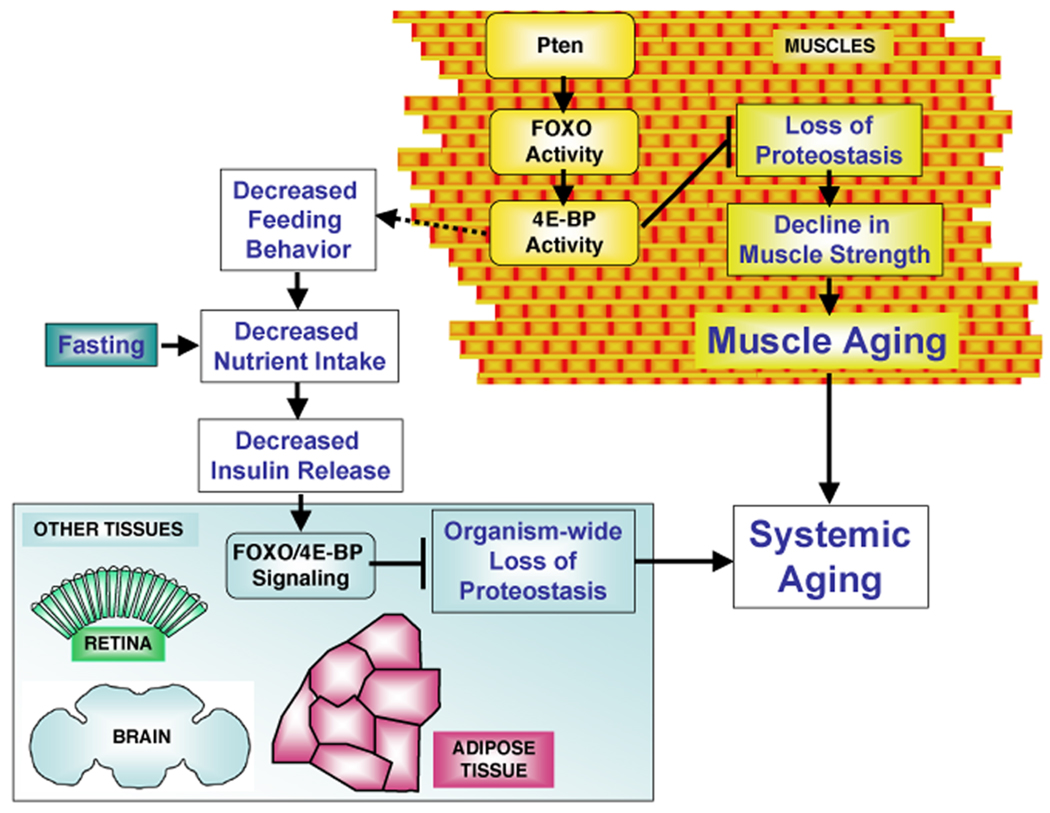
The progressive loss of muscle strength during aging is a common degenerative event of unclear pathogenesis. Although muscle functional decline precedes age-related changes in other tissues, its contribution to systemic aging is unknown. Here, we show that muscle aging is characterized in Drosophila by the progressive accumulation of protein aggregates that associate with impaired muscle function. The transcription factor FOXO and its target 4E-BP remove damaged proteins at least in part via the autophagy/lysosome system, whereas foxo mutants have dysfunctional proteostasis. Both FOXO and 4E-BP delay muscle functional decay and extend life span. Moreover, FOXO/4E-BP signaling in muscles decreases feeding behavior and the release of insulin from producing cells, which in turn delays the age-related accumulation of protein aggregates in other tissues. These findings reveal an organism-wide regulation of proteostasis in response to muscle aging and a key role of FOXO/4E-BP signaling in the coordination of organismal and tissue aging.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
References
-
- Arndt V, Dick N, Tawo R, Dreiseidler M, Wenzel D, Hesse M, Furst DO, Saftig P, Saint R, Fleischmann BK, et al. Chaperone-Assisted Selective Autophagy Is Essential for Muscle Maintenance. Curr Biol. 2010;20:143–148. - PubMed
-
- Augustin H, Partridge L. Invertebrate Models of Age-Related Muscle Degeneration. Biochim Biophys Acta. 2009;1790:1084–1094. - PubMed
-
- Bodine SC, Stitt TN, Gonzalez M, Kline WO, Stover GL, Bauerlein R, Zlotchenko E, Scrimgeour A, Lawrence JC, Glass DJ, Yancopoulos GD. Akt/mTOR pathway is a crucial regulator of skeletal muscle hypertrophy and can prevent muscle atrophy in vivo. Nat Cell Biol. 2001;3:1014–1019. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
