

Protein folding in the cell: challenges and progress
- PMID: 21112769
- PMCID: PMC3072030
- DOI: 10.1016/j.sbi.2010.11.001
Protein folding in the cell: challenges and progress
Abstract
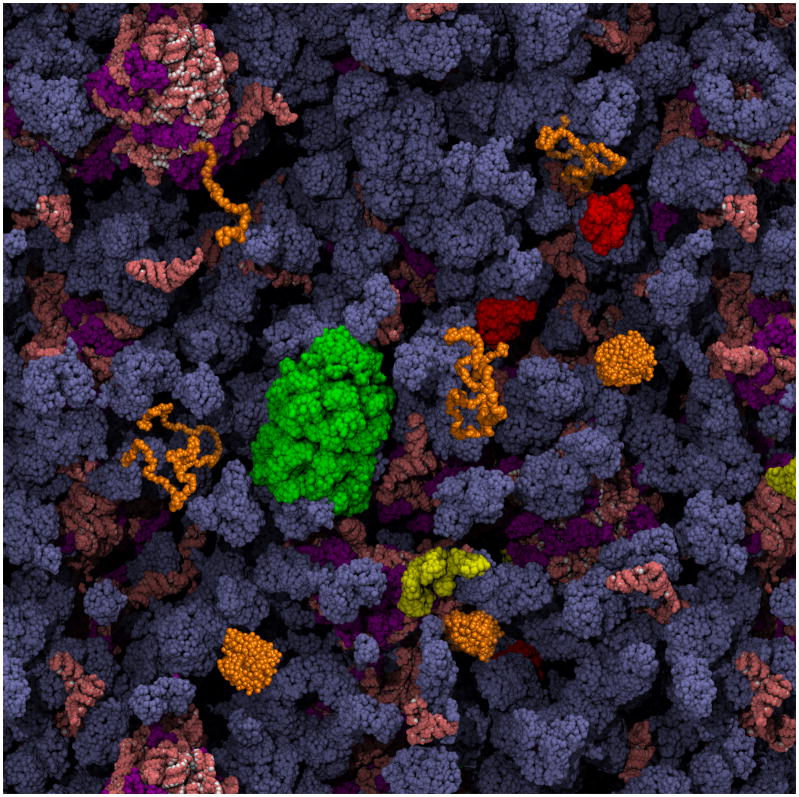
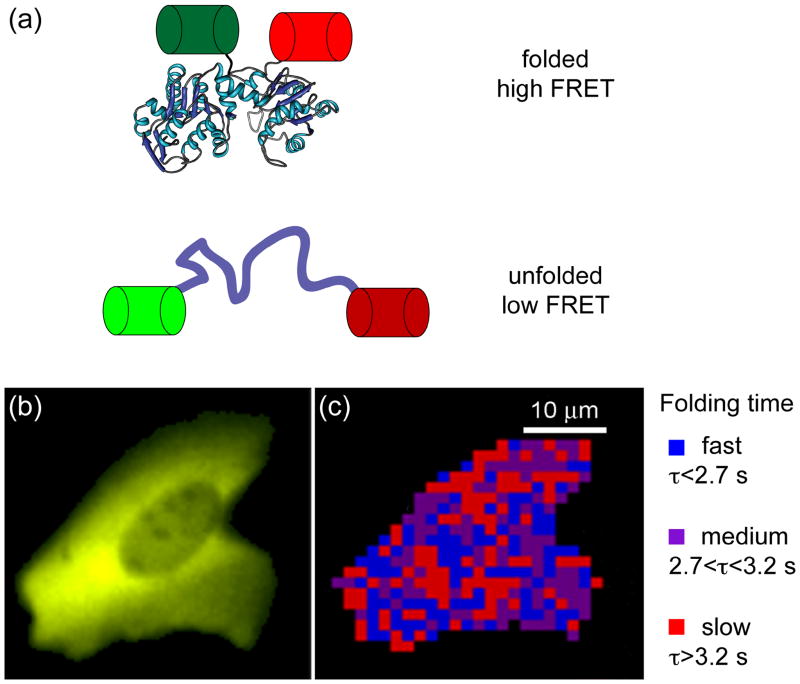
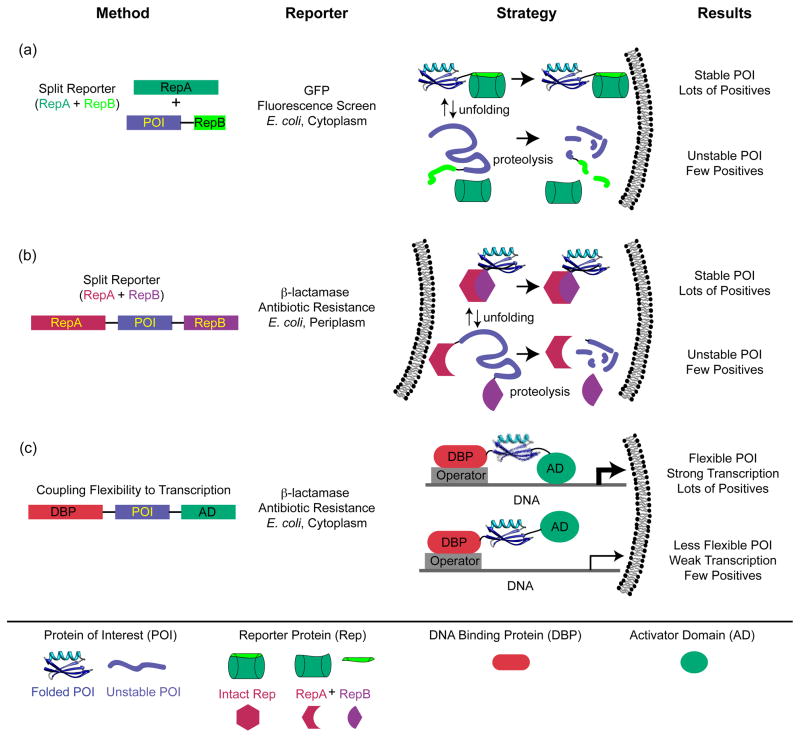
It is hard to imagine a more extreme contrast than that between the dilute solutions used for in vitro studies of protein folding and the crowded, compartmentalized, sticky, spatially inhomogeneous interior of a cell. This review highlights recent research exploring protein folding in the cell with a focus on issues that are generally not relevant to in vitro studies of protein folding, such as macromolecular crowding, hindered diffusion, cotranslational folding, molecular chaperones, and evolutionary pressures. The technical obstacles that must be overcome to characterize protein folding in the cell are driving methodological advances, and we draw attention to several examples, such as fluorescence imaging of folding in cells and genetic screens for in-cell stability.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Wang Y, He H, Li S. Effect of Ficoll 70 on thermal stability and structure of creatine kinase. Biochemistry. 2010;75:648–654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
