

Reversing EphB2 depletion rescues cognitive functions in Alzheimer model
- PMID: 21113149
- PMCID: PMC3030448
- DOI: 10.1038/nature09635
Reversing EphB2 depletion rescues cognitive functions in Alzheimer model
Abstract
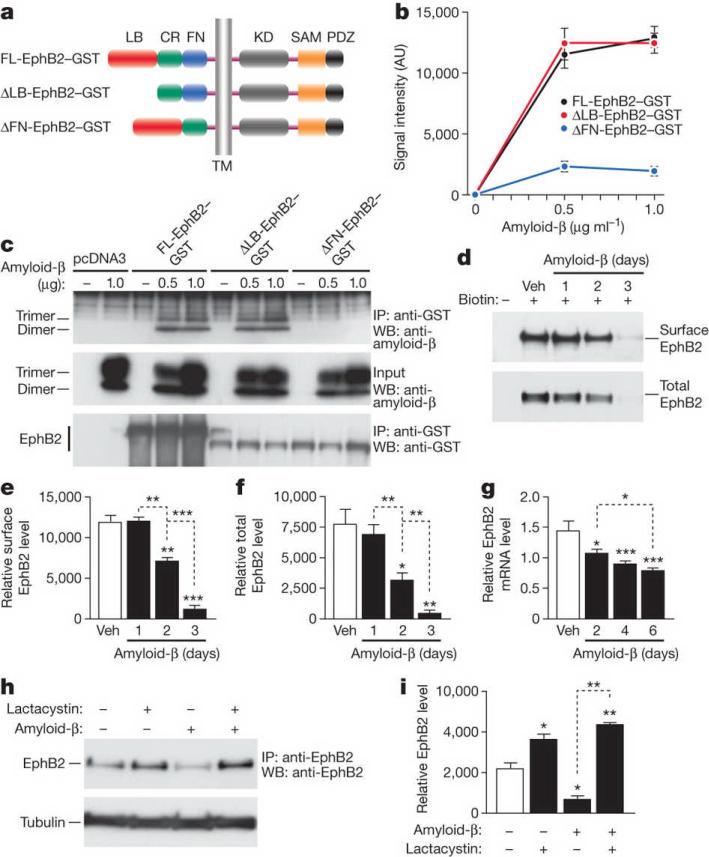
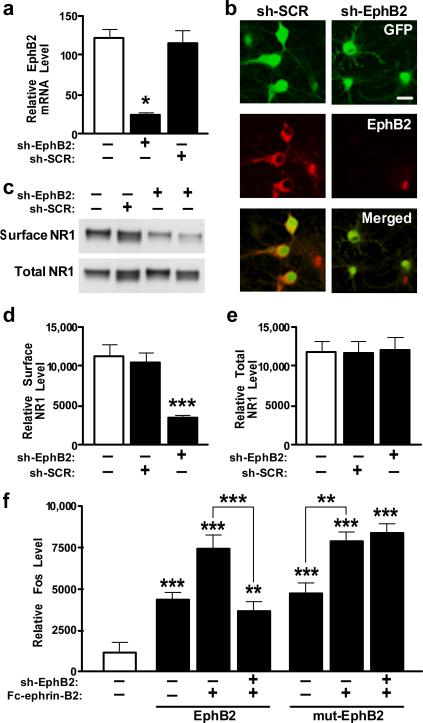
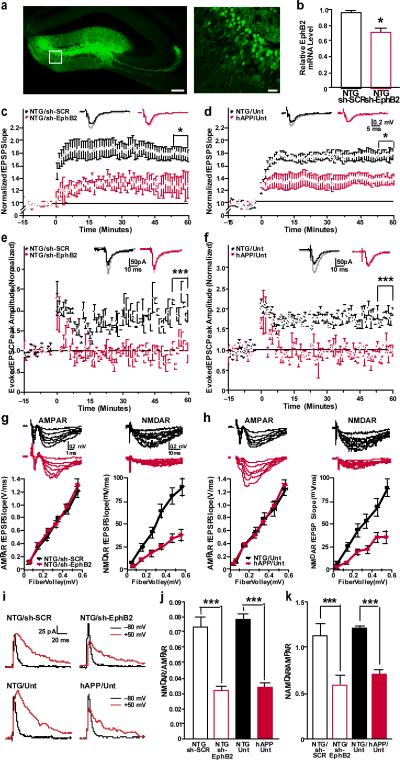
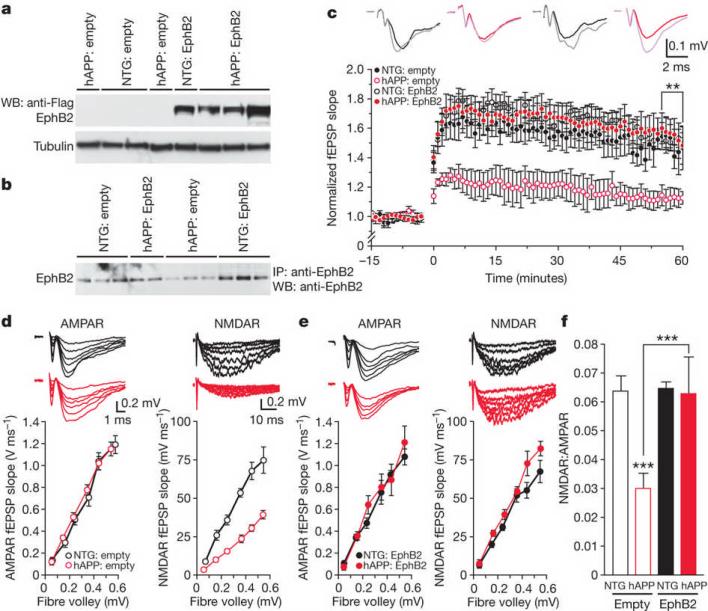
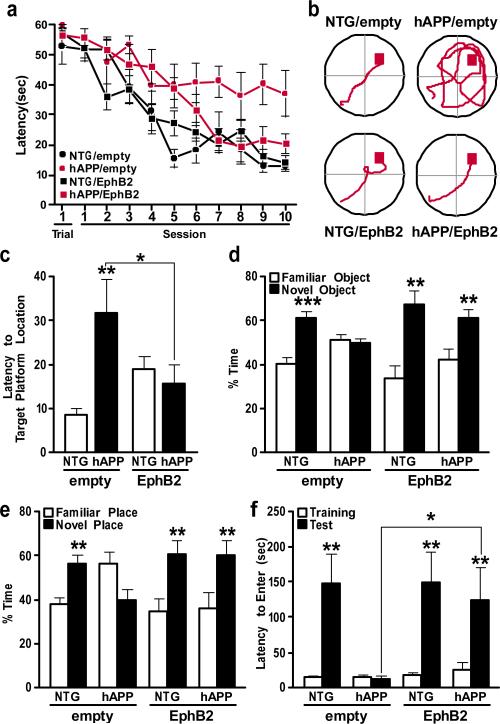
Amyloid-β oligomers may cause cognitive deficits in Alzheimer's disease by impairing neuronal NMDA-type glutamate receptors, whose function is regulated by the receptor tyrosine kinase EphB2. Here we show that amyloid-β oligomers bind to the fibronectin repeats domain of EphB2 and trigger EphB2 degradation in the proteasome. To determine the pathogenic importance of EphB2 depletions in Alzheimer's disease and related models, we used lentiviral constructs to reduce or increase neuronal expression of EphB2 in memory centres of the mouse brain. In nontransgenic mice, knockdown of EphB2 mediated by short hairpin RNA reduced NMDA receptor currents and impaired long-term potentiation in the dentate gyrus, which are important for memory formation. Increasing EphB2 expression in the dentate gyrus of human amyloid precursor protein transgenic mice reversed deficits in NMDA receptor-dependent long-term potentiation and memory impairments. Thus, depletion of EphB2 is critical in amyloid-β-induced neuronal dysfunction. Increasing EphB2 levels or function could be beneficial in Alzheimer's disease.
Figures
Comment in
-
Alzheimer's disease: Recollection of lost memories.Nature. 2011 Jan 6;469(7328):44-5. doi: 10.1038/469044a. Nature. 2011. PMID: 21209657 Free PMC article.
-
Alzheimer disease: EphB2 depletion links amyloid-β to cognitive impairment.Nat Rev Neurol. 2011 Feb;7(2):62. doi: 10.1038/nrneurol.2010.204. Nat Rev Neurol. 2011. PMID: 21391318 No abstract available.
References
-
- Walsh DM, Selkoe DJ. Deciphering the molecular basis of memory failure in Alzheimer's disease. Neuron. 2004;44:181–193. - PubMed
-
- Kamenetz F, et al. APP processing and synaptic function. Neuron. 2003;37:925–937. - PubMed
-
- Malenka RC, Bear MF. LTP and LTD: an embarrassment of riches. Neuron. 2004;44:5–21. - PubMed
-
- Ikonomovic MD, et al. Distribution of glutamate receptor subunit NMDAR1 in the hippocampus of normal elderly and patients with Alzheimer's disease. Exp. Neurol. 1999;160:194–204. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
