

Effects of cholesterol depletion on compartmentalized cAMP responses in adult cardiac myocytes
- PMID: 21115018
- PMCID: PMC3049871
- DOI: 10.1016/j.yjmcc.2010.11.015
Effects of cholesterol depletion on compartmentalized cAMP responses in adult cardiac myocytes
Abstract
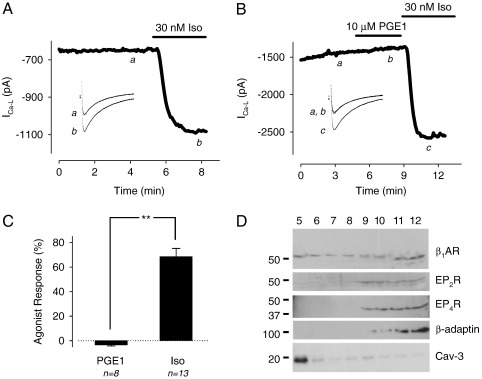
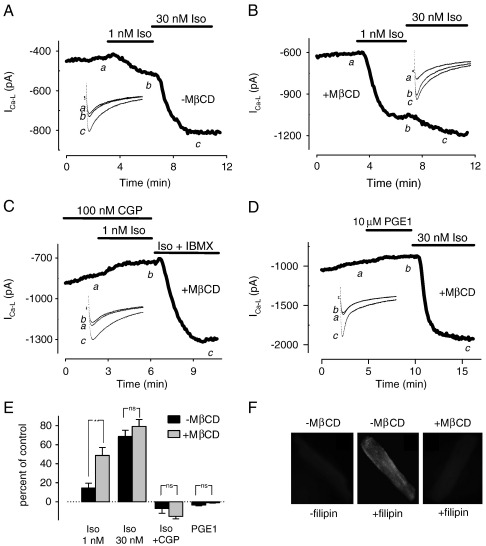
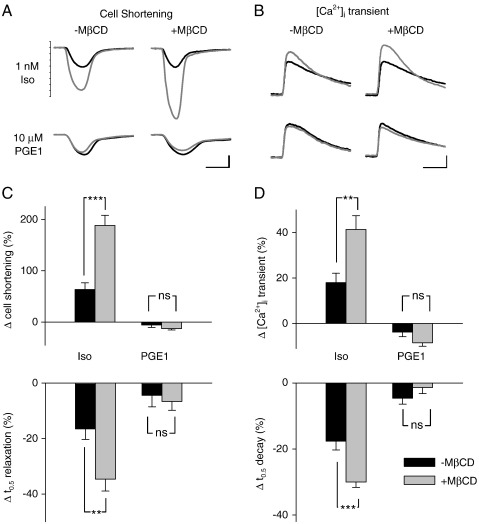
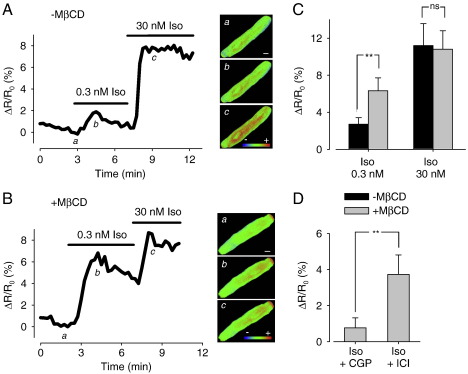
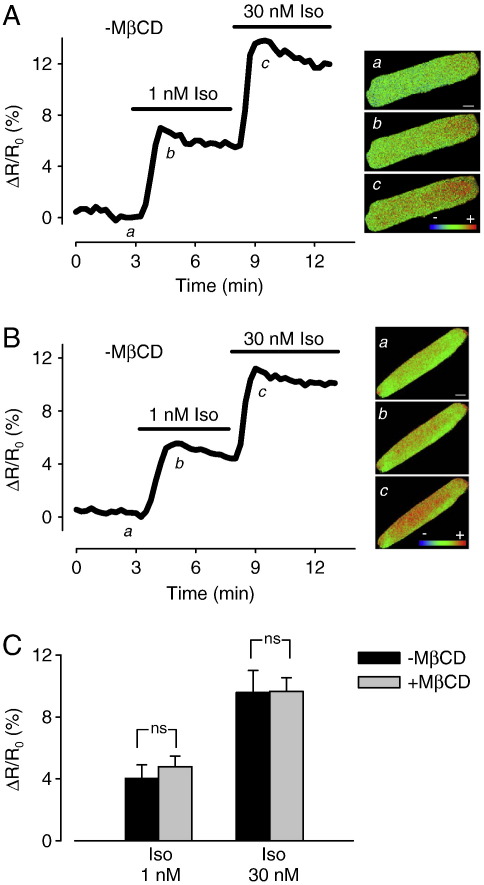
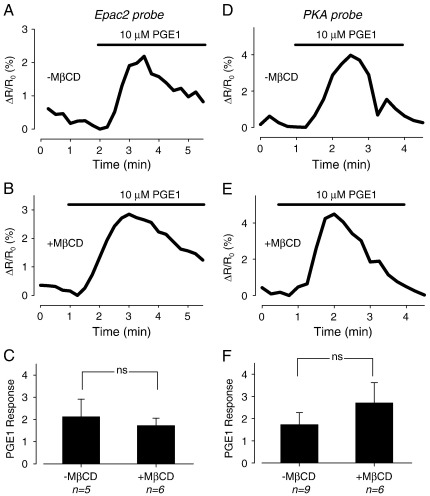
β(1)-Adrenergic receptors (β(1)ARs) and E-type prostaglandin receptors (EPRs) both produce compartmentalized cAMP responses in cardiac myocytes. The role of cholesterol-dependent lipid rafts in producing these compartmentalized responses was investigated in adult rat ventricular myocytes. β(1)ARs were found in lipid raft and non-lipid raft containing membrane fractions, while EPRs were only found in non-lipid raft fractions. Furthermore, β(1)AR activation enhanced the L-type Ca(2+) current, intracellular Ca(2+) transient, and myocyte shortening, while EPR activation had no effect, consistent with the idea that these functional responses are regulated by cAMP produced by receptors found in lipid raft domains. Using methyl-β-cyclodextrin to disrupt lipid rafts by depleting membrane cholesterol did not eliminate compartmentalized behavior, but it did selectively alter specific receptor-mediated responses. Cholesterol depletion enhanced the sensitivity of functional responses produced by β(1)ARs without having any effect on EPR activation. Changes in cAMP activity were also measured in intact cells using two different FRET-based biosensors: a type II PKA-based probe to monitor cAMP in subcellular compartments that include microdomains associated with caveolar lipid rafts and a freely diffusible Epac2-based probe to monitor total cytosolic cAMP. β(1)AR and EPR activation elicited responses detected by both FRET probes. However, cholesterol depletion only affected β(1)AR responses detected by the PKA probe. These results indicate that lipid rafts alone are not sufficient to explain the difference between β(1)AR and EPR responses. They also suggest that β(1)AR regulation of myocyte contraction involves the local production of cAMP by a subpopulation of receptors associated with caveolar lipid rafts.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Hartzell H.C. Regulation of cardiac ion channels by catecholamines, acetylcholine and second messenger systems. Prog. Biophys. Molec. Biol. 1988;52:165–247. - PubMed
-
- Bers D.M. Kluwer; Dordrecht: 2001. Excitation–contraction coupling and cardiac contractile force.
-
- Buxton I.L., Brunton L.L. Compartments of cyclic AMP and protein kinase in mammalian cardiomyocytes. J. Biol. Chem. 1983;258:10233–10239. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
