

Telomeres in cancer and ageing
- PMID: 21115533
- PMCID: PMC3001316
- DOI: 10.1098/rstb.2010.0291
Telomeres in cancer and ageing
Abstract
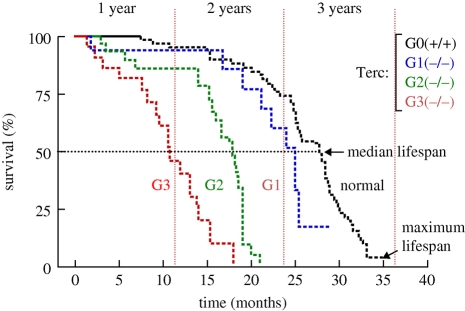
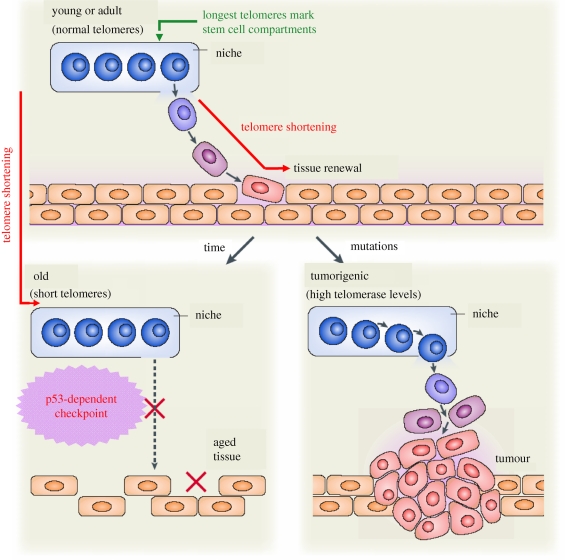
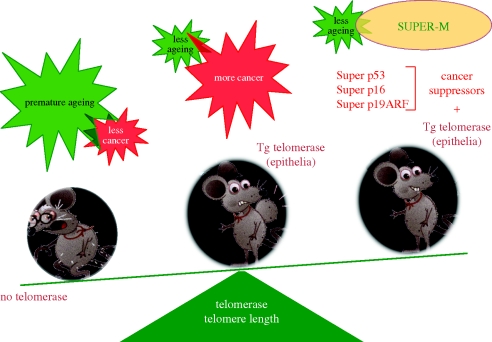
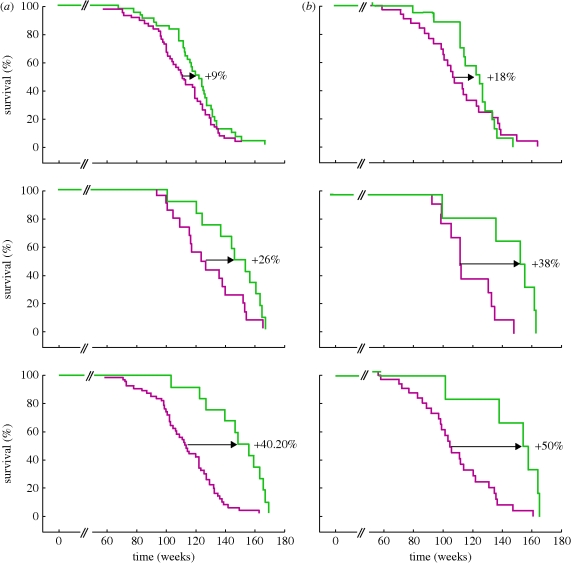
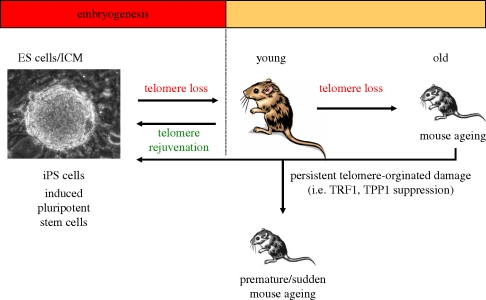
Telomeres protect the chromosome ends from unscheduled DNA repair and degradation. Telomeres are heterochromatic domains composed of repetitive DNA (TTAGGG repeats) bound to an array of specialized proteins. The length of telomere repeats and the integrity of telomere-binding proteins are both important for telomere protection. Furthermore, telomere length and integrity are regulated by a number of epigenetic modifications, thus pointing to higher order control of telomere function. In this regard, we have recently discovered that telomeres are transcribed generating long, non-coding RNAs, which remain associated with the telomeric chromatin and are likely to have important roles in telomere regulation. In the past, we showed that telomere length and the catalytic component of telomerase, Tert, are critical determinants for the mobilization of stem cells. These effects of telomerase and telomere length on stem cell behaviour anticipate the premature ageing and cancer phenotypes of telomerase mutant mice. Recently, we have demonstrated the anti-ageing activity of telomerase by forcing telomerase expression in mice with augmented cancer resistance. Shelterin is the major protein complex bound to mammalian telomeres; however, its potential relevance for cancer and ageing remained unaddressed to date. To this end, we have generated mice conditionally deleted for the shelterin proteins TRF1, TPP1 and Rap1. The study of these mice demonstrates that telomere dysfunction, even if telomeres are of a normal length, is sufficient to produce premature tissue degeneration, acquisition of chromosomal aberrations and initiation of neoplastic lesions. These new mouse models, together with the telomerase-deficient mouse model, are valuable tools for understanding human pathologies produced by telomere dysfunction.
Figures
References
-
- de Lange T.2005Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev. 19, 2100–2110 10.1101/gad.1346005 (doi:10.1101/gad.1346005) - DOI - PubMed
-
- Blasco M. A.2007The epigenetic regulation of mammalian telomeres. Nat. Rev. Genet. 8, 299–309 10.1038/nrg2047 (doi:10.1038/nrg2047) - DOI - PubMed
-
- Greider C. W., Blackburn E. H.1985Identification of a specific telomere terminal transferase activity in Tetrahymena extracts. Cell 43, 405–413 10.1016/0092-8674(85)90170-9 (doi:10.1016/0092-8674(85)90170-9) - DOI - PubMed
-
- Blasco M. A.2005Telomeres and human disease: aging, cancer and beyond. Nat. Rev. Genet. 6, 611–622 10.1038/nrg1656 (doi:10.1038/nrg1656) - DOI - PubMed
-
- Armanios M. Y., et al. 2007Telomerase mutations in families with idiopathic pulmonary fibrosis. N. Engl. J. Med. 356, 1317–1326 10.1056/NEJMoa066157 (doi:10.1056/NEJMoa066157) - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous