

Visualization of glutamate as a volume transmitter
- PMID: 21115644
- PMCID: PMC3055537
- DOI: 10.1113/jphysiol.2010.199539
Visualization of glutamate as a volume transmitter
Abstract
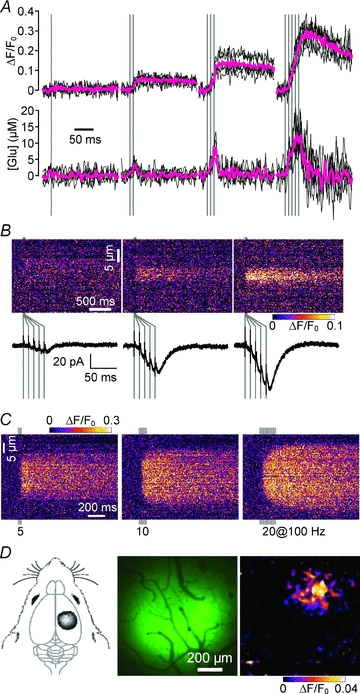
Glutamate is the major excitatory neurotransmitter in the central nervous system. Although glutamate mediates synaptically confined point-to-point transmission, it has been suggested that under certain conditions glutamate may escape from the synaptic cleft (glutamate spillover), accumulate in the extrasynaptic space, and mediate volume transmission to regulate important brain functions. However, the inability to directly measure glutamate dynamics around active synapses has limited our understanding of glutamatergic volume transmission. The recent development of a family of fluorescent glutamate indicators has enabled the visualization of extrasynaptic glutamate dynamics in brain tissues. In this topical review, we examine glutamate as a volume transmitter based on novel results of glutamate imaging in the brain.
Figures


References
-
- Agulhon C, Fiacco TA, McCarthy KD. Hippocampal short- and long-term plasticity are not modulated by astrocyte Ca2+ signaling. Science. 2010;327:1250–1254. - PubMed
-
- Arnth-Jensen N, Jabaudon D, Scanziani M. Cooperation between independent hippocampal synapses is controlled by glutamate uptake. Nat Neurosci. 2002;5:325–331. - PubMed
-
- Asztely F, Erdemli G, Kullmann DM. Extrasynaptic glutamate spillover in the hippocampus: dependence on temperature and the role of active glutamate uptake. Neuron. 1997;18:281–293. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources