

FtsZ in bacterial cytokinesis: cytoskeleton and force generator all in one
- PMID: 21119015
- PMCID: PMC3008173
- DOI: 10.1128/MMBR.00021-10
FtsZ in bacterial cytokinesis: cytoskeleton and force generator all in one
Abstract
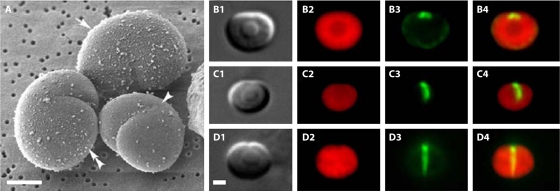
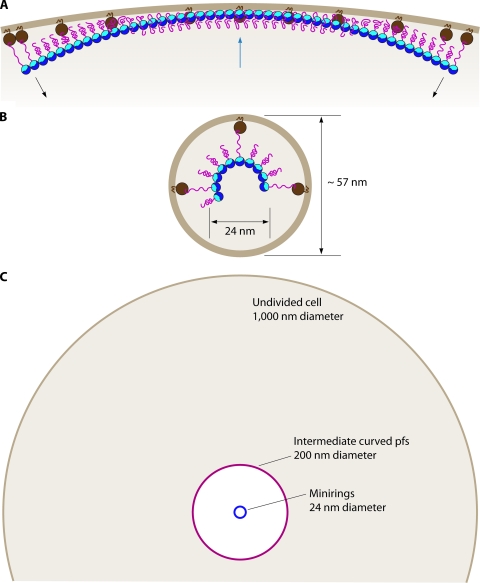
FtsZ, a bacterial homolog of tubulin, is well established as forming the cytoskeletal framework for the cytokinetic ring. Recent work has shown that purified FtsZ, in the absence of any other division proteins, can assemble Z rings when incorporated inside tubular liposomes. Moreover, these artificial Z rings can generate a constriction force, demonstrating that FtsZ is its own force generator. Here we review light microscope observations of how Z rings assemble in bacteria. Assembly begins with long-pitch helices that condense into the Z ring. Once formed, the Z ring can transition to short-pitch helices that are suggestive of its structure. FtsZ assembles in vitro into short protofilaments that are ∼30 subunits long. We present models for how these protofilaments might be further assembled into the Z ring. We discuss recent experiments on assembly dynamics of FtsZ in vitro, with particular attention to how two regulatory proteins, SulA and MinC, inhibit assembly. Recent efforts to develop antibacterial drugs that target FtsZ are reviewed. Finally, we discuss evidence of how FtsZ generates a constriction force: by protofilament bending into a curved conformation.
Figures



References
-
- Aarsman, M. E., A. Piette, C. Fraipont, T. M. Vinkenvleugel, M. Nguyen-Disteche, and T. den Blaauwen. 2005. Maturation of the Escherichia coli divisome occurs in two steps. Mol. Microbiol. 55:1631-1645. - PubMed
-
- Adams, D. W., and J. Errington. 2009. Bacterial cell division: assembly, maintenance and disassembly of the Z ring. Nat. Rev. Microbiol. 7:642-653. - PubMed
-
- Addinall, S. G., C. Cao, and J. Lutkenhaus. 1997. Temperature shift experiments with ftsZ84(Ts) strain reveal rapid dynamics of FtsZ localization and indicate that the Z ring is required throughout septation and cannot reoccupy division sites once constriction has initiated. J. Bacteriol. 179:4277-4284. - PMC - PubMed
-
- Addinall, S. G., and J. Lutkenhaus. 1996. FtsZ-spirals and -arcs determine the shape of the invaginating septa in some mutants of Escherichia coli. Mol. Microbiol. 22:231-237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
