

B-function expression in the flower center underlies the homeotic phenotype of Lacandonia schismatica (Triuridaceae)
- PMID: 21119062
- PMCID: PMC3015125
- DOI: 10.1105/tpc.109.069153
B-function expression in the flower center underlies the homeotic phenotype of Lacandonia schismatica (Triuridaceae)
Erratum in
- Plant Cell. 2011 Jan;23(1):428. Vergara-Silva, Francisco [removed]
Abstract
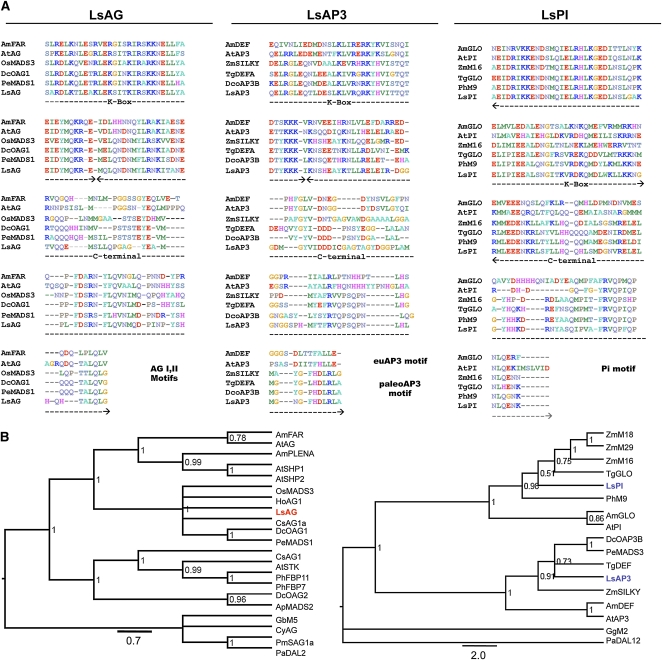
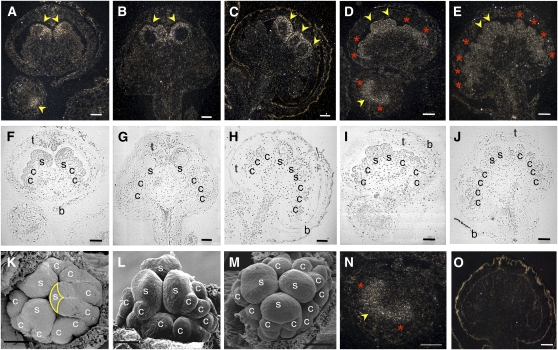
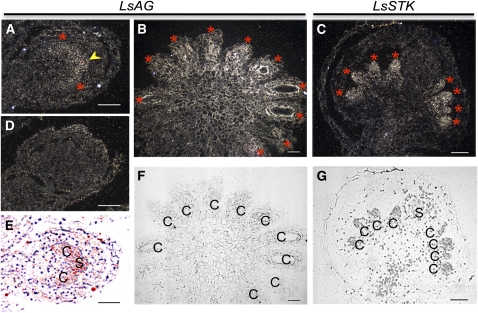
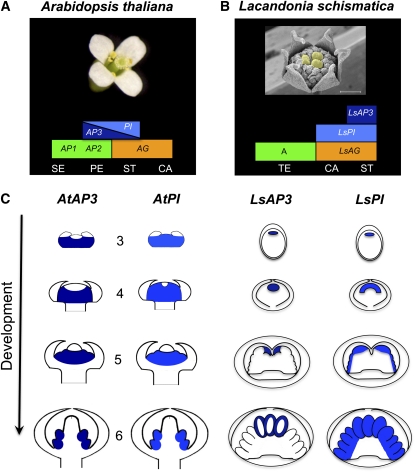
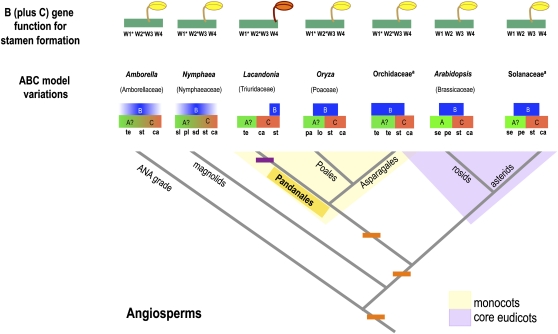
Spontaneous homeotic transformations have been described in natural populations of both plants and animals, but little is known about the molecular-genetic mechanisms underlying these processes in plants. In the ABC model of floral organ identity in Arabidopsis thaliana, the B- and C-functions are necessary for stamen morphogenesis, and C alone is required for carpel identity. We provide ABC model-based molecular-genetic evidence that explains the unique inside-out homeotic floral organ arrangement of the monocotyledonous mycoheterotroph species Lacandonia schismatica (Triuridaceae) from Mexico. Whereas a quarter million flowering plant species bear central carpels surrounded by stamens, L. schismatica stamens occur in the center of the flower and are surrounded by carpels. The simplest explanation for this is that the B-function is displaced toward the flower center. Our analyses of the spatio-temporal pattern of B- and C-function gene expression are consistent with this hypothesis. The hypothesis is further supported by conservation between the B-function genes of L. schismatica and Arabidopsis, as the former are able to rescue stamens in Arabidopsis transgenic complementation lines, and Ls-AP3 and Ls-PI are able to interact with each other and with the corresponding Arabidopsis B-function proteins in yeast. Thus, relatively simple molecular modifications may underlie important morphological shifts in natural populations of extant plant taxa.
Figures

References
-
- Álvarez-Buylla E.R., Azpeitia E., Barrio R., Benítez M., Padilla-Longoria P. (2010a). From ABC genes to regulatory networks, epigenetic landscapes and flower morphogenesis: making biological sense of theoretical approaches. Semin. Cell Dev. Biol. 21: 108–117 - PubMed
-
- Álvarez-Buylla E.R., Benítez M., Corvera-Poiré A., Chaos-Cador A., de Folter S., Gamboa de Buen A., Garay-Arroyo A., García-Ponce B., Jaimes-Miranda F., Pérez-Ruiz R.V., Piñeyro-Nelson A., Sánchez-Corrales Y.E. (March 23, 2010b). Flower development. In The Arabidopsis Book, Somerville C.R., Meyerowitz E.M., (Rockville, MD: American Society of Plant Biologists; ), doi/10.1199/tab.0127, http://www.aspb.org/publications/arabidopsis/ - PMC - PubMed
-
- Ambrose B.A., Espinosa-Matías S., Vázquez-Santana S., Vergara-Silva F., Martínez E., Márquez-Guzmán J., Álvarez-Buylla E.R. (2006). Comparative developmental series of the Mexican triurids support a euanthial interpretation for the unusual reproductive axes of Lacandonia schismatica (Triuridaceae). Am. J. Bot. 93: 15–35
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
