

Coupling governs entrainment range of circadian clocks
- PMID: 21119632
- PMCID: PMC3010105
- DOI: 10.1038/msb.2010.92
Coupling governs entrainment range of circadian clocks
Abstract
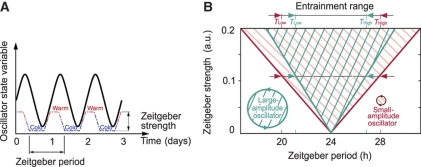
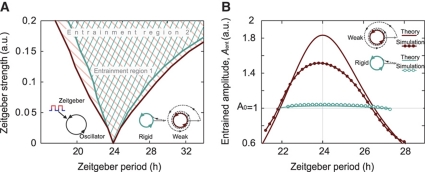
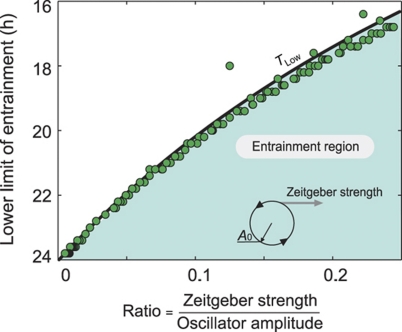
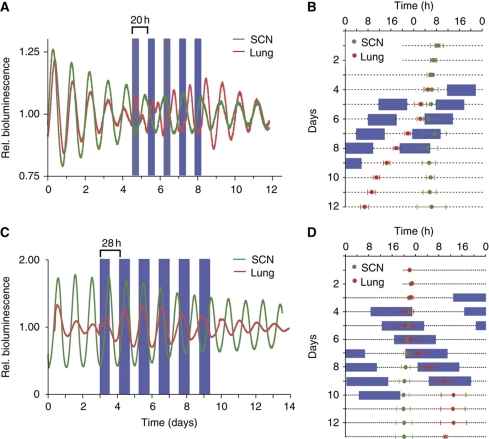
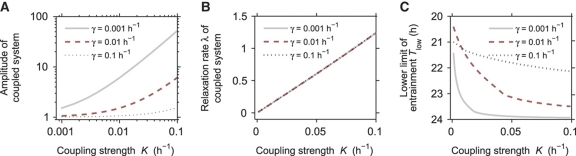
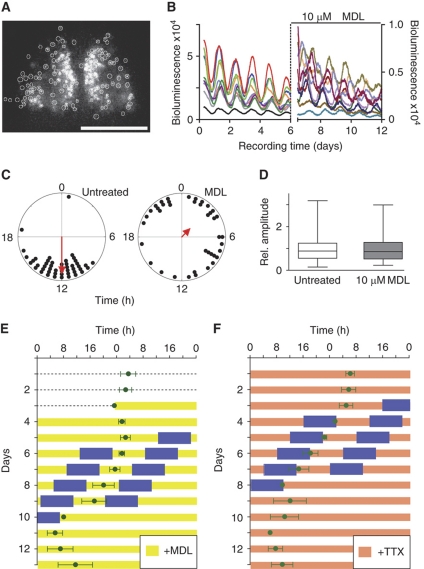
Circadian clocks are endogenous oscillators driving daily rhythms in physiology and behavior. Synchronization of these timers to environmental light-dark cycles ('entrainment') is crucial for an organism's fitness. Little is known about which oscillator qualities determine entrainment, i.e., entrainment range, phase and amplitude. In a systematic theoretical and experimental study, we uncovered these qualities for circadian oscillators in the suprachiasmatic nucleus (SCN-the master clock in mammals) and the lung (a peripheral clock): (i) the ratio between stimulus (zeitgeber) strength and oscillator amplitude and (ii) the rigidity of the oscillatory system (relaxation rate upon perturbation) determine entrainment properties. Coupling among oscillators affects both qualities resulting in increased amplitude and rigidity. These principles explain our experimental findings that lung clocks entrain to extreme zeitgeber cycles, whereas SCN clocks do not. We confirmed our theoretical predictions by showing that pharmacological inhibition of coupling in the SCN leads to larger ranges of entrainment. These differences between master and the peripheral clocks suggest that coupling-induced rigidity in the SCN filters environmental noise to create a robust circadian system.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Amir S, Cain S, Sullivan J, Robinson B, Stewart J (1999) Olfactory stimulation enhances light-induced phase shifts in free-running activity rhythms and Fos expression in the suprachiasmatic nucleus. Neuroscience 92: 1165–1170 - PubMed
-
- Anishchenko VS, Astakhov V, Neiman A, Vadivasova T, Schimansky-Geier L (2007) Nonlinear Dynamics of Chaotic and Stochastic Systems: Tutorial and Modern Developments. Springer-Verlag New York: LLC
-
- Aschoff J, Pohl H (1978) Phase relations between a circadian rhythm and its zeitgeber within the range of entrainment. Naturwissenschaften 65: 80–84 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
