

Conservation, loss, and redeployment of Wnt ligands in protostomes: implications for understanding the evolution of segment formation
- PMID: 21122121
- PMCID: PMC3003278
- DOI: 10.1186/1471-2148-10-374
Conservation, loss, and redeployment of Wnt ligands in protostomes: implications for understanding the evolution of segment formation
Abstract
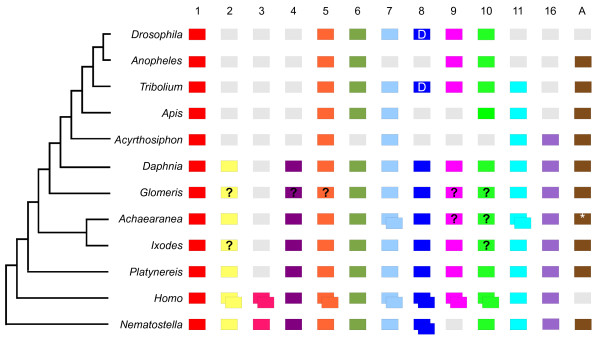
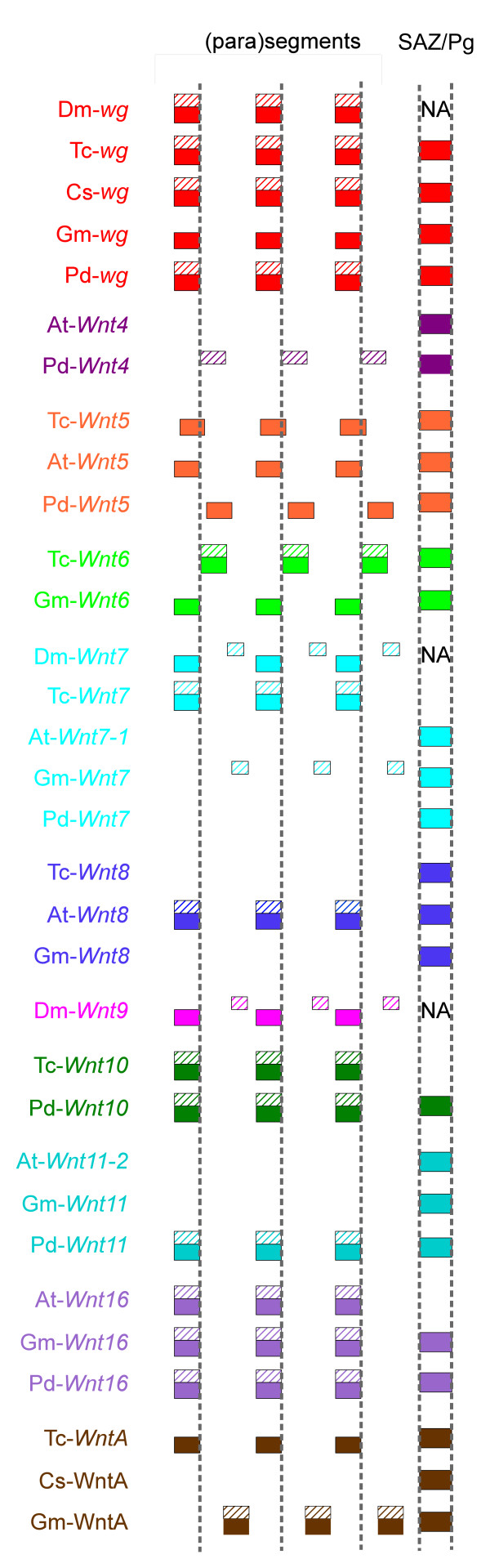
Background: The Wnt genes encode secreted glycoprotein ligands that regulate a wide range of developmental processes, including axis elongation and segmentation. There are thirteen subfamilies of Wnt genes in metazoans and this gene diversity appeared early in animal evolution. The loss of Wnt subfamilies appears to be common in insects, but little is known about the Wnt repertoire in other arthropods, and moreover the expression and function of these genes have only been investigated in a few protostomes outside the relatively Wnt-poor model species Drosophila melanogaster and Caenorhabditis elegans. To investigate the evolution of this important gene family more broadly in protostomes, we surveyed the Wnt gene diversity in the crustacean Daphnia pulex, the chelicerates Ixodes scapularis and Achaearanea tepidariorum, the myriapod Glomeris marginata and the annelid Platynereis dumerilii. We also characterised Wnt gene expression in the latter three species, and further investigated expression of these genes in the beetle Tribolium castaneum.
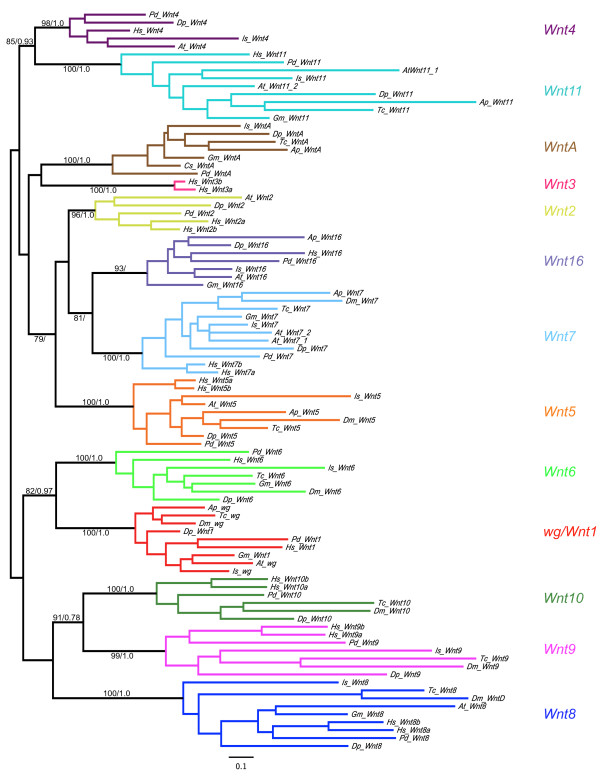
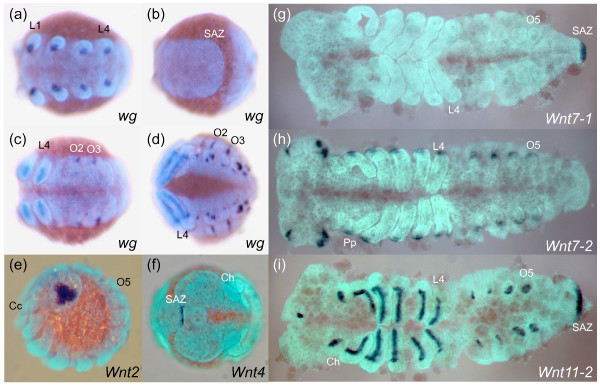
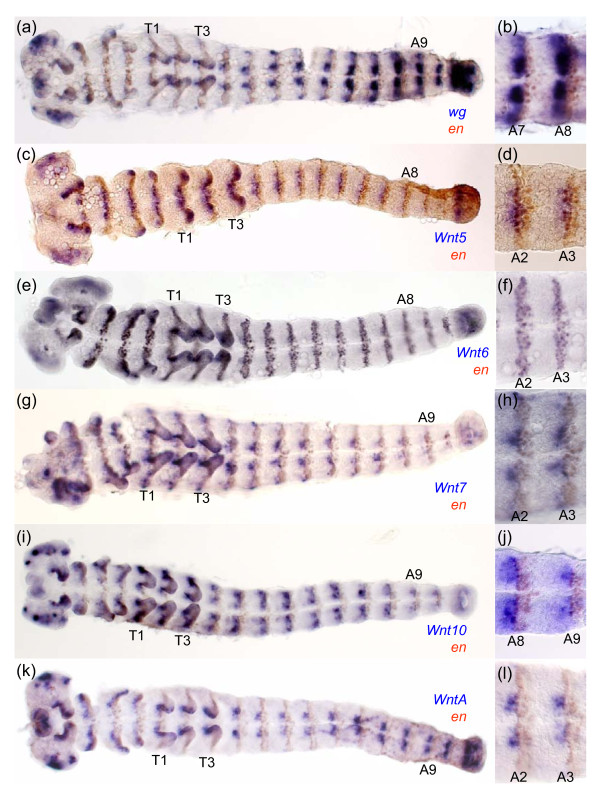
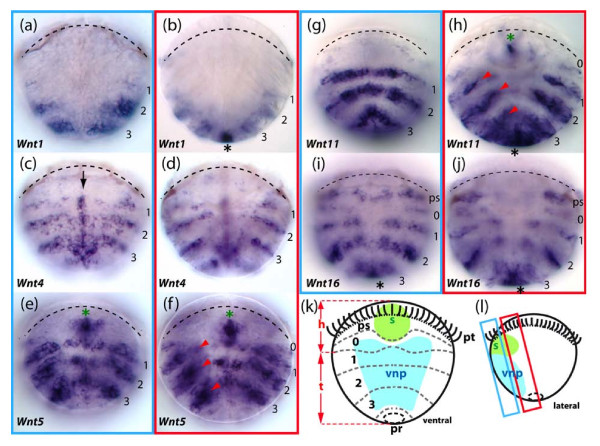
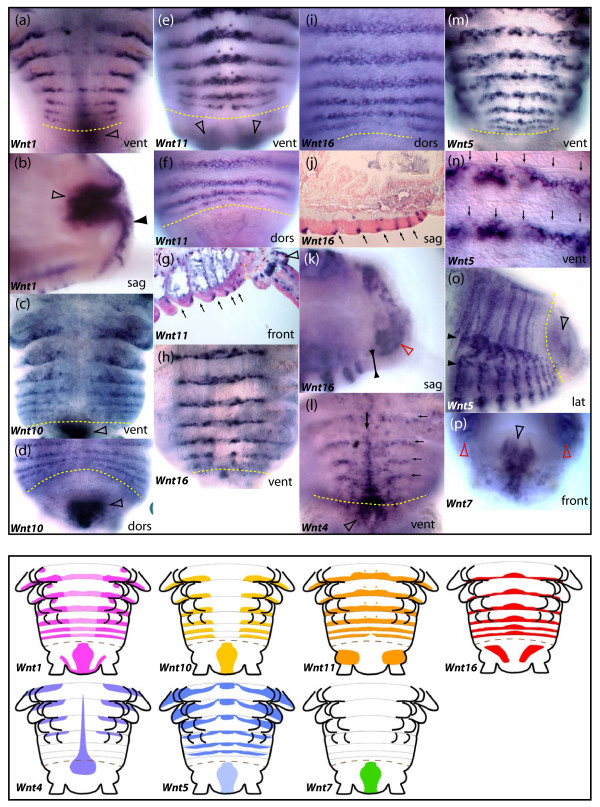
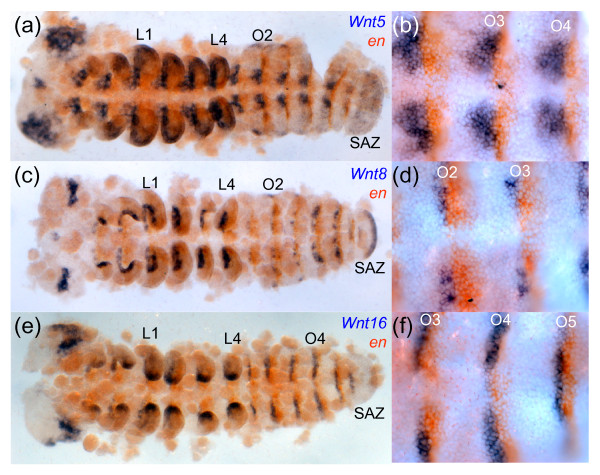
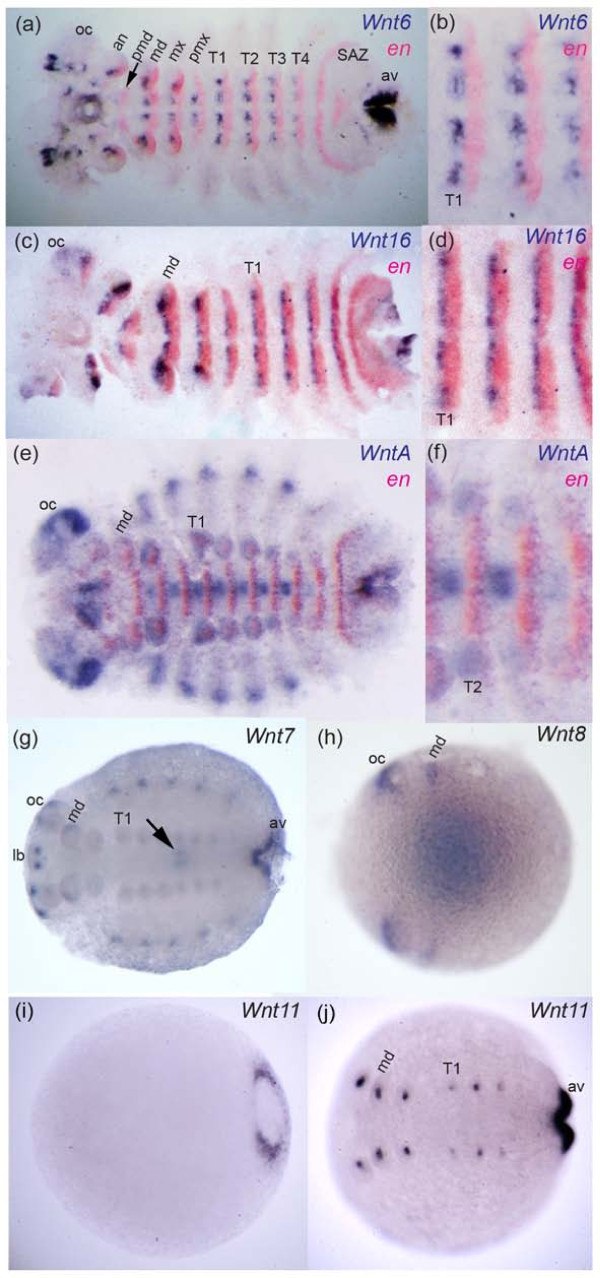
Results: We found that Daphnia and Platynereis both contain twelve Wnt subfamilies demonstrating that the common ancestors of arthropods, ecdysozoans and protostomes possessed all members of all Wnt subfamilies except Wnt3. Furthermore, although there is striking loss of Wnt genes in insects, other arthropods have maintained greater Wnt gene diversity. The expression of many Wnt genes overlap in segmentally reiterated patterns and in the segment addition zone, and while these patterns can be relatively conserved among arthropods and the annelid, there have also been changes in the expression of some Wnt genes in the course of protostome evolution. Nevertheless, our results strongly support the parasegment as the primary segmental unit in arthropods, and suggest further similarities between segmental and parasegmental regulation by Wnt genes in annelids and arthropods respectively.
Conclusions: Despite frequent losses of Wnt gene subfamilies in lineages such as insects, nematodes and leeches, most protostomes have probably maintained much of their ancestral repertoire of twelve Wnt genes. The maintenance of a large set of these ligands could be in part due to their combinatorial activity in various tissues rather than functional redundancy. The activity of such Wnt 'landscapes' as opposed to the function of individual ligands could explain the patterns of conservation and redeployment of these genes in important developmental processes across metazoans. This requires further analysis of the expression and function of these genes in a wider range of taxa.
Figures
References
-
- Graba Y, Gieseler K, Aragnol D, Laurenti P, Mariol MC, Berenger H, Sagnier T, Pradel J. DWnt-4, a novel Drosophila Wnt gene acts downstream of homeotic complex genes in the visceral mesoderm. Development. 1995;121(1):209–218. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
