

Gait-specific energetics contributes to economical walking and running in emus and ostriches
- PMID: 21123267
- PMCID: PMC3107644
- DOI: 10.1098/rspb.2010.2022
Gait-specific energetics contributes to economical walking and running in emus and ostriches
Abstract
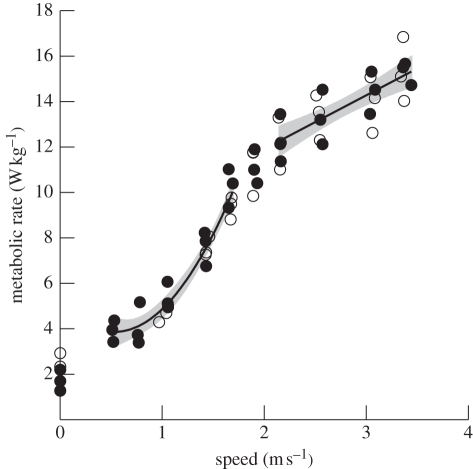
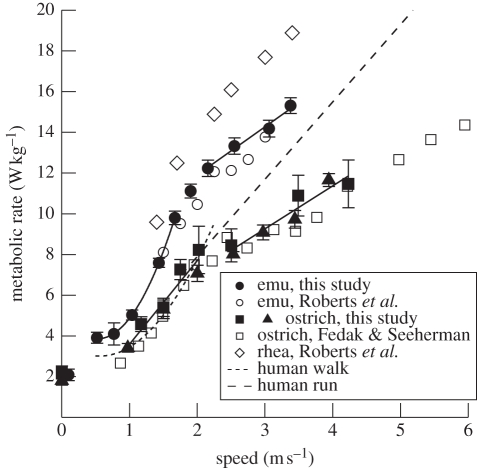
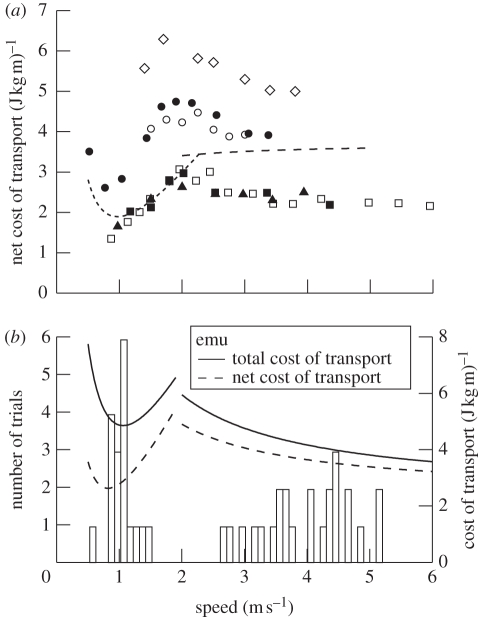
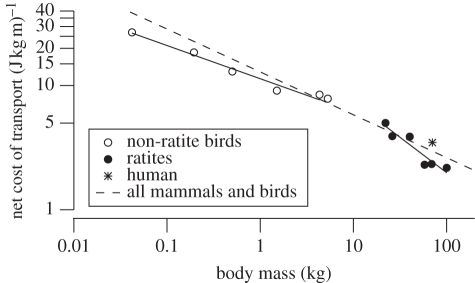
A widely held assumption is that metabolic rate (Ė(met)) during legged locomotion is linked to the mechanics of different gaits and this linkage helps explain the preferred speeds of animals in nature. However, despite several prominent exceptions, Ė(met) of walking and running vertebrates has been nearly uniformly characterized as increasing linearly with speed across all gaits. This description of locomotor energetics does not predict energetically optimal speeds for minimal cost of transport (E(cot)). We tested whether large bipedal ratite birds (emus and ostriches) have gait-specific energetics during walking and running similar to those found in humans. We found that during locomotion, emus showed a curvilinear relationship between Ė(met) and speed during walking, and both emus and ostriches demonstrated an abrupt change in the slope of Ė(met) versus speed at the gait transition with a linear increase during running. Similar to human locomotion, the minimum net E(cot) calculated after subtracting resting metabolism was lower in walking than in running in both species. However, the difference in net E(cot) between walking and running was less than is found in humans because of a greater change in the slope of Ė(met) versus speed at the gait transition, which lowers the cost of running for the avian bipeds. For emus, we also show that animals moving freely overground avoid a range of speeds surrounding the gait-transition speed within which the E(cot) is large. These data suggest that deviations from a linear relation of metabolic rate and speed and variations in transport costs with speed are more widespread than is often assumed, and provide new evidence that locomotor energetics influences the choice of speed in bipedal animals. The low cost of transport for walking is probably ecologically important for emus and ostriches because they spend the majority of their active day walking, and thus the energy used for locomotion is a large part of their daily energy budget.
Figures
Similar articles
-
Preferred gait and walk-run transition speeds in ostriches measured using GPS-IMU sensors.J Exp Biol. 2016 Oct 15;219(Pt 20):3301-3308. doi: 10.1242/jeb.142588. J Exp Biol. 2016. PMID: 27802152
-
Gait selection in the ostrich: mechanical and metabolic characteristics of walking and running with and without an aerial phase.Proc Biol Sci. 2004 May 22;271(1543):1091-9. doi: 10.1098/rspb.2004.2702. Proc Biol Sci. 2004. PMID: 15293864 Free PMC article.
-
Gait-specific metabolic costs and preferred speeds in ring-tailed lemurs (Lemur catta), with implications for the scaling of locomotor costs.Am J Phys Anthropol. 2012 Nov;149(3):356-64. doi: 10.1002/ajpa.22132. Epub 2012 Sep 14. Am J Phys Anthropol. 2012. PMID: 22976581
-
Scaling of avian bipedal locomotion reveals independent effects of body mass and leg posture on gait.J Exp Biol. 2018 May 22;221(Pt 10):jeb152538. doi: 10.1242/jeb.152538. J Exp Biol. 2018. PMID: 29789347 Review.
-
Patterns of mechanical energy change in tetrapod gait: pendula, springs and work.J Exp Zool A Comp Exp Biol. 2006 Nov 1;305(11):899-911. doi: 10.1002/jez.a.334. J Exp Zool A Comp Exp Biol. 2006. PMID: 17029267 Review.
Cited by
-
A new approach to quantify semiochemical effects on insects based on energy landscapes.PLoS One. 2014 Aug 29;9(8):e106276. doi: 10.1371/journal.pone.0106276. eCollection 2014. PLoS One. 2014. PMID: 25171062 Free PMC article.
-
Economical Speed and Energetically Optimal Transition Speed Evaluated by Gross and Net Oxygen Cost of Transport at Different Gradients.PLoS One. 2015 Sep 18;10(9):e0138154. doi: 10.1371/journal.pone.0138154. eCollection 2015. PLoS One. 2015. PMID: 26383249 Free PMC article. Clinical Trial.
-
Study on bio-inspired feet based on the cushioning and shock absorption characteristics of the ostrich foot.PLoS One. 2020 Jul 24;15(7):e0236324. doi: 10.1371/journal.pone.0236324. eCollection 2020. PLoS One. 2020. PMID: 32706841 Free PMC article.
-
Sex differences in gait utilization and energy metabolism during terrestrial locomotion in two varieties of chicken (Gallus gallus domesticus) selected for different body size.Biol Open. 2015 Sep 24;4(10):1306-15. doi: 10.1242/bio.013094. Biol Open. 2015. PMID: 26405047 Free PMC article.
-
Natural Frequency Method: estimating the preferred walking speed of Tyrannosaurus rex based on tail natural frequency.R Soc Open Sci. 2021 Apr 21;8(4):201441. doi: 10.1098/rsos.201441. R Soc Open Sci. 2021. PMID: 33996115 Free PMC article.
References
-
- Pennycuick C. J. 1975. On the running of the gnu (Connochaetes taurinus) and other animals. J. Exp. Biol. 63, 775–799
-
- Hoyt D. F., Taylor C. R. 1981. Gait and the energetics of locomotion in horses. Nature 292, 239–24010.1038/292239a0 (doi:10.1038/292239a0) - DOI - DOI
-
- Perry A. K., Blickhan R., Biewener A. A., Heglund N. C., Taylor C. R. 1988. Preferred speeds in terrestrial vertebrates: are they equivalent? J. Exp. Biol. 137, 207–219 - PubMed
-
- Wickler S. J., Hoyt D. F., Cogger E. A., Hall K. M. 2001. Effect of load on preferred speed and cost of transport. J. Appl. Physiol. 90, 1548–1551 - PubMed
-
- Alexander R. M. 1989. Optimization and gaits in the locomotion of vertebrates. Physiol. Rev. 69, 1199–1227 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
