

doi: 10.1128/JVI.02134-10.
Epub 2010 Dec 1.
The adenovirus E1b55K/E4orf6 complex induces degradation of the Bloom helicase during infection
Affiliations
- PMID: 21123383
- PMCID: PMC3028918
- DOI: 10.1128/JVI.02134-10
Item in Clipboard
The adenovirus E1b55K/E4orf6 complex induces degradation of the Bloom helicase during infection
J Virol.
2011 Feb.
Abstract
The adenovirus (Ad) E1b55K and E4orf6 gene products assemble an E3 ubiquitin ligase complex that promotes degradation of cellular proteins. Among the known substrates are p53 and the Mre11-Rad50-Nbs1 (MRN) complex. Since members of the RecQ helicase family function together with MRN in genome maintenance, we investigated whether adenovirus affects RecQ proteins. We show that Bloom helicase (BLM) is degraded during adenovirus type 5 (Ad5) infection. BLM degradation is mediated by E1b55K/E4orf6 but is independent of MRN. We detected BLM localized at discrete foci around viral replication centers. These studies identify BLM as a new substrate for degradation by the adenovirus E1b55K/E4orf6 complex.
Figures
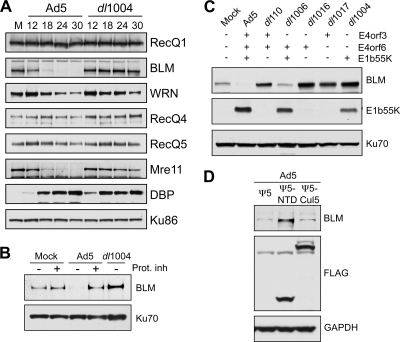
Adenovirus infection induces proteasome-mediated degradation of BLM. (A) The steady-state levels of cellular RecQ helicases were examined over a time course of adenovirus infection. HeLa cells were either mock infected (M) or infected with wild-type Ad5 (multiplicity of infection [MOI] of 10) or the E4-deleted mutant dl1004 (MOI of 25). Cells were harvested at the indicated hours postinfection (hpi) for analysis by immunoblotting (14). Specific antibodies were used to detect RecQ1 (Santa Cruz), WRN (BD Biosciences), RecQ4 (Cell Signaling), and RecQ5 (gift from P. Janscak) (32). To generate the anti-BLM antibody, rabbits were immunized with a purified recombinant protein consisting of a His-tagged BLM fragment (amino acid residues 1 to 439). The purified anti-BLM antiserum (designated 7099) was tested for specificity by immunoblotting and immunofluorescence (data not shown). The viral DBP (detected with monoclonal antibody B6, from A. Levine) served as a control for infection, and degradation of cellular proteins was confirmed by inclusion of the Mre11 positive control (Genetex). Antibody to Ku86 (Santa Cruz) served as a loading control. Although the levels of WRN were slightly affected, only BLM was reduced in an E4-dependent manner analogous to Mre11 degradation. (B) BLM degradation is proteasome dependent. Cells were infected with Ad5 (MOI of 10) or dl1004 (MOI of 25), and at 12 hpi, proteasome inhibitors (Prot. inh) (10 μM MG132 and 1 μM epoxomicin) were added to the cells for a further 12 h. Degradation of BLM by Ad5 was prevented by the proteasome inhibitors. Ku70 served as a loading control. (C) E1b55K and E4orf6 are required for degradation of BLM during adenovirus infection. BLM levels were compared in HeLa cells infected for 24 h with wild-type Ad5 or mutants lacking genes from the E1 and E4 regions as indicated (2, 10, 11, 26). Compared with results for mock-infected cells, BLM levels were reduced only during infection with viruses that express both E1b55K and E4orf6. E1b55K (detected with monoclonal antibody 2A6, from A. Levine) and Ku70 (antibody from Santa Cruz) served as controls for infection and gel loading, respectively. (D) The Cul5 complex is required for BLM degradation. Cells were infected with Ad5 (MOI of 10), superinfected (MOI of 50) with Ψ5, Ψ5-Cul5, or Ψ5-NTD (68), and harvested at 24 h after the primary infection. In these infections, the FLAG antibody (Sigma) demonstrates expression of Cul5 or NTD-Cul5 from the Ψ5 viruses, and GAPDH (glyceraldehyde-3-phosphate dehydrogenase) (Research Diagnostics Inc.) serves as a loading control.
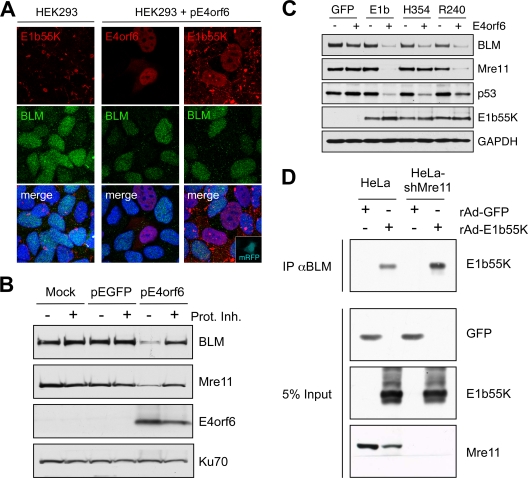
E1b55K and E4orf6 are sufficient for degradation of BLM. (A) Immunofluorescence reveals that in HEK293 cells the BLM protein is located throughout the nucleoplasm and E1b55K is in cytoplasmic aggregates (left). When HEK293 cells were transfected with E4orf6 (15), BLM levels were reduced (middle). In cells transfected with plasmids expressing E4orf6 and monomeric red fluorescent protein (mRFP) (at a 9:1 ratio), transfected cells demonstrated nuclear E1b55K, with reduced BLM levels (right). Cells were fixed for staining (14) at 24 h after transfection, and DAPI (4′,6-diamidino-2-phenylindole) staining indicates the location of the nuclei in all merged images. (B) Expression of E4orf6 by transfection of HEK293 cells demonstrated a proteasome-dependent decrease in BLM levels. Cells were harvested at 20 h posttransfection for analysis by immunoblotting with specific antibodies. Degradation was abrogated by proteasome inhibitors (10 μM MG132 and 1 μM epoxomicin). Mre11 served as a control for degradation, and Ku70 served as a loading control. (C) E1b55K and E4orf6 are sufficient for BLM degradation. E4orf6 was expressed by rAd vector transduction (48) of U2OS cells that stably express GFP or E1b55K (14), and protein levels were assessed by immunoblotting with the indicated antibodies. Antibodies to Mre11 (Genetex) and p53 (Calbiochem) served as controls for degradation, and Ku70 served as a loading control. (D) E1b55K coimmunoprecipitates with BLM in the absence of Mre11. HeLa cells or HeLa-shMre11 cells were infected (MOI of 50) with rAd-GFP and rAd-E1b55K (48) for 24 h, and lysates were subjected to immunoprecipitation with the BLM antibody (IP αBLM). Immunoblotting of the precipitated proteins demonstrated that E1b55K could be pulled down by BLM in lysates from both cells.
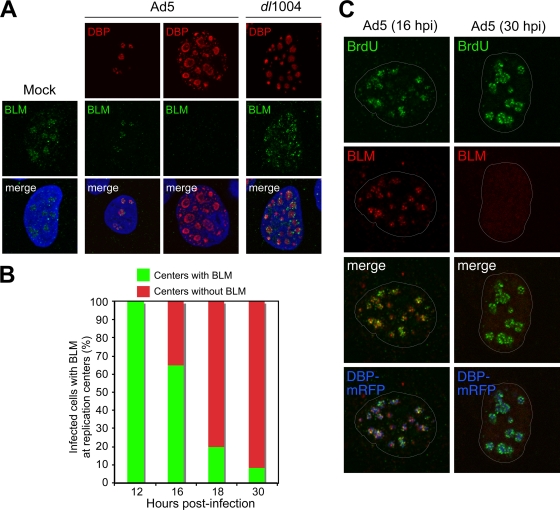
Accumulation of BLM at sites of viral replication. (A) HeLa cells were infected with wild-type Ad5 (MOI of 10) or E4 mutant dl1004 (MOI of 25). Localization of BLM was examined by immunofluorescence (14) after preextraction to remove soluble protein prior to fixation and compared to that for mock-infected cells (left). In uninfected cells, BLM localized to nucleoli and ND10 structures. In Ad5-infected cells, BLM was either located in discrete foci surrounding early viral replication centers (as detected with an antibody to the viral DBP) or undetectable in cells with large centers (representing late-stage infection). In cells infected with E4-deleted virus dl1004, all infected cells displayed BLM accumulated at viral centers. DAPI staining indicates the location of the nuclei in all merged images. (B) Quantification of infected cells with BLM at viral replication centers over a time course of wild-type Ad5 infection. At each time point, the relative number of cells with BLM in the two patterns demonstrated by the representative images shown in panel A was determined by examining 100 infected cells. (C) BLM at sites of active viral replication. HeLa cells were transfected with DBP-mRFP and, after 8 h, were infected with Ad5 (MOI of 10). At early (16 hpi) and late (30 hpi) stages of infection, cells were pulsed with BrdU for 30 min to label sites of DNA replication (46) and detected with a BrdU-specific antibody (Sigma). In preextracted cells, BLM could be detected adjacent to sites of ongoing viral replication. By 30 hpi, BLM had been degraded.
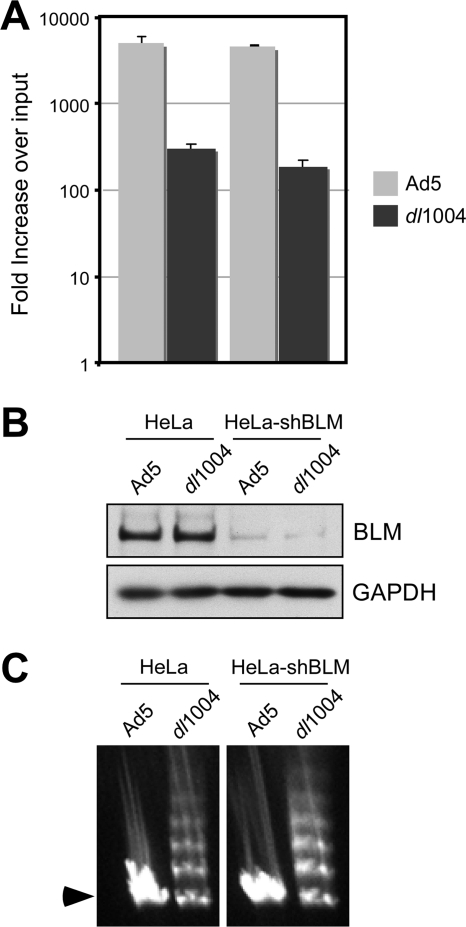
BLM is not responsible for defective viral replication or concatemer formation. (A) Viral replication was assessed in HeLa cells and HeLa cells stably expressing shRNA against BLM (HeLa-shBLM cells). The shRNA sequence 5′-TGCCAATGACCAGGCGATC was inserted into the pSUPER.retro.puro vector (Orbigen), and Phoenix retroviral packaging cells were used to produce retrovirus. HeLa cells were transduced with retrovirus, and the HeLa-shBLM cells were selected and maintained in puromycin. Cells were infected with Ad5 (MOI of 10) or dl1004 (MOI of 3) in triplicate, and DNA was extracted for quantitative PCR at 4 and 30 hpi as previously described (35). Accumulation of viral DNA is represented as fold increase over input viral DNA, as determined at the 4-h time point, and error bars represent standard errors of the means (SEM) from the triplicate samples. (B) Immunoblotting of lysates from infected cells at the 4-h time point confirmed knockdown of BLM levels in HeLa-shBLM cells. GAPDH served as a loading control. (C) Concatemer formation was examined by PFGE analysis of DNA at 48 hpi with Ad5 (MOI of 25) and dl1004 (MOI of 50) as previously described (57). Viral DNA was visualized by staining the gel in ethidium bromide, and the position of the linear viral genome is indicated by an arrowhead. Formation of concatemers by dl1004 was not dependent on BLM.
References
-
- Blanchette, P., and P. E. Branton. 2009. Manipulation of the ubiquitin-proteasome pathway by small DNA tumor viruses. Virology 384:317-323. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
