

Review
doi: 10.1101/cshperspect.a004390.
Protein folding in the cytoplasm and the heat shock response
Affiliations
- PMID: 21123396
- PMCID: PMC2982175
- DOI: 10.1101/cshperspect.a004390
Item in Clipboard
Review
Protein folding in the cytoplasm and the heat shock response
Cold Spring Harb Perspect Biol.
2010 Dec.
Abstract
Proteins generally must fold into precise three-dimensional conformations to fulfill their biological functions. In the cell, this fundamental process is aided by molecular chaperones, which act in preventing protein misfolding and aggregation. How this machinery assists newly synthesized polypeptide chains in navigating the complex folding energy landscape is now being understood in considerable detail. The mechanisms that ensure the maintenance of a functional proteome under normal and stress conditions are also of great medical relevance, as the aggregation of proteins that escape the cellular quality control underlies a range of debilitating diseases, including many age-of-onset neurodegenerative disorders.
Figures
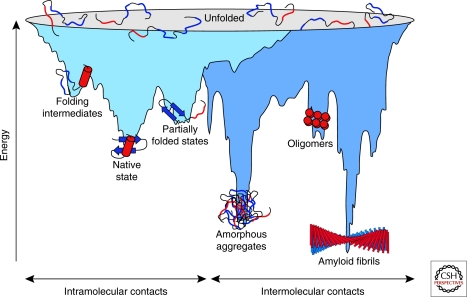
Energy landscape of protein folding and aggregation. The light blue surface shows the multitude of conformations “funneling” to the native state via intramolecular contacts and the dark blue area the conformations moving toward amorphous aggregates or amyloid fibrils via intermolecular contacts. Both parts of the energy surface overlap. Aggregate formation can occur from intermediates populated during de novo folding or by destabilization of the native state and is normally prevented by molecular chaperones. Cell-toxic oligomers may occur as off-pathway intermediates of amyloid fibril formation. (Reprinted, with permission of Macmillan Publishers Ltd., from Hartl and Hayer-Hartl 2009.)
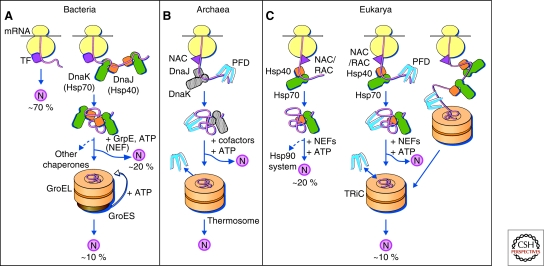
Protein folding in the cytosol. (A) Models for the chaperone-assisted folding of newly synthesized polypeptides in the cytosol. Bacteria (left panel). TF, trigger factor; N, native protein. Nascent chains probably interact generally with TF, and most small proteins (∼70% of total) may fold rapidly on synthesis without further assistance. Longer chains interact subsequently with DnaK and DnaJ (Hsp70 system) and fold on one or several cycles of ATP-dependent binding and release (∼20% of total). About 10% of chains transit the chaperonin system (GroEL and GroES) for folding. (B) Archaea (middle panel). PFD, prefoldin; NAC, nascent chain-associated complex. Note that only some archaeal species contain DnaK/DnaJ. (C) Eukarya (right panel). Like TF, NAC (nascent chain associated complex) probably interacts generally with nascent chains, but the role of NAC in folding is not yet clear. About 20% of chains reach their native states in a reaction assisted by RAC (ribosome associated complex), Hsp70, and Hsp40. A fraction of these must be transferred to Hsp90 for folding. About 10% of chains are co- or posttranslationally passed on to the chaperonin TRiC in a reaction mediated by Hsp70 and PFD, both of which interact directly with TRiC. PFD recognizes the nascent chains of certain TRiC substrates, including actin and tubulins. (Modified, with permission of The American Association for the Advancement of Science, from Hartl and Hayer-Hartl 2002.)
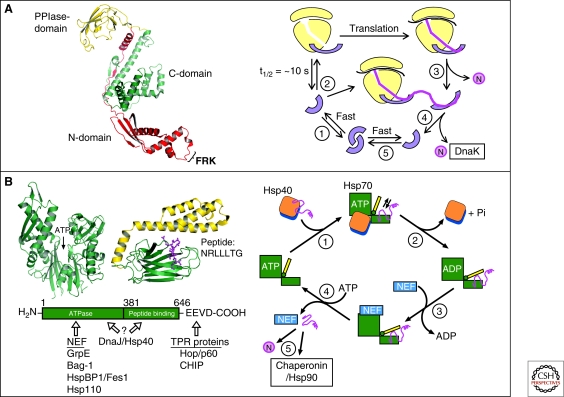
Chaperone paradigms: Trigger factor and Hsp70 system. (A) Trigger factor (TF): The left panel shows the domain structure of TF, including the FRK ribosome-binding loop in the amino-domain (PDB 1W26) (Ferbitz et al. 2004), and the right panel shows a model of the TF reaction cycle (modified from Kaiser et al. [2006]). (1) Free TF is in rapid equilibrium between monomeric and dimeric states. (2) TF monomer binds to nontranslating ribosomes with a KD of ∼1 µM and a mean residence time of approximately 10 seconds. Ribosome binding causes a conformational expansion of TF and may activate TF for interaction with nascent chains. (3) Nascent chains that interact weakly with TF may begin to compact cotranslationally in the vicinity of TF. Release of TF from nascent chain coincides with TF dissociation from the ribosome and allows completion of folding to native state (N). (4) Structurally more complex proteins may strongly interact with TF. TF remains bound to nascent chain after dissociating from the ribosome and a new TF molecule can enter at the ribosome. Eventual chain dissociation from TF facilitates transfer to DnaK or folding. (5) Released TF enters the monomer-dimer pool. (B) Hsp70 system. Left panel: Structures of the ATPase domain (PDB 1DKG) and the peptide-binding domain (PDB 1DKZ) (Zhu et al. 1996) of Hsp70 shown representatively for Escherichia coli DnaK. The α-helical lid of the peptide binding domain is shown in yellow and the extended peptide substrate as a ball-and-stick model in pink. ATP indicates the position of the nucleotide binding site. The amino acid sequence of the peptide is indicated in single-letter code. The interaction of prokaryotic and eukaryotic cofactors with Hsp70 is shown schematically. Residue numbers refer to human Hsp70. Only the Hsp70 proteins of the eukaryotic cytosol have the COOH-terminal sequence EEVD that is involved in binding of tetratricopeptide repeat (TPR) cofactors (Scheufler et al. 2000). Right panel: Hsp70 reaction cycle. NEF, nucleotide exchange factor (GrpE in case of E. coli DnaK; Bag, HspBP1, and Hsp110 in case of eukaryotic cytosolic Hsp70). N, native protein. (A,B: Modified with permission from Hartl and Hayer-Hartl 2009.)
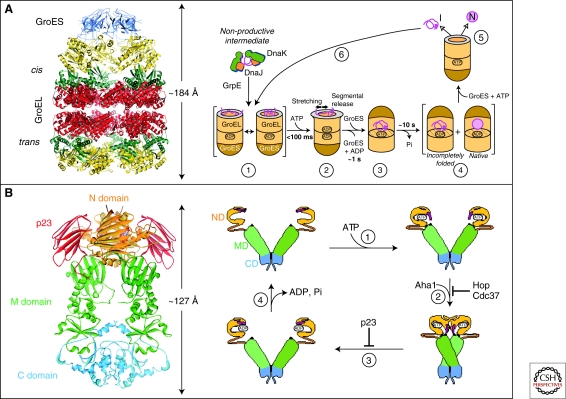
Chaperone paradigms: Chaperonins and Hsp90 system. (A) The GroEL-GroES chaperonin. Left panel: Crystal structure of the asymmetric GroEL-GroES complex (PDB 1AON) (Xu et al. 1997). Cis, the GroES-bound chamber of GroEL. Trans, the opposite GroEL ring. Right panel: Working model summarizing the conformational changes in a substrate protein on transfer from DnaK/DnaJ (Hsp70 system) to GroEL and during GroEL/GroES-mediated folding. Note that binding of a second substrate molecule to the open ring of GroEL in steps 4 and 5 as well as the transient formation of a symmetrical GroEL-GroES2 complex is omitted for simplicity. (1) Substrate protein may be delivered to GroEL by DnaK/DnaJ in a nonaggregated, but kinetically trapped state. On binding to GroEL it undergoes local unfolding to an ensemble of expanded and more compact conformations. (2) ATP-dependent domain movement of the apical GroEL domains result in stretching of tightly bound regions of substrate and in release and partial compaction of less stably bound regions. (3) Compaction is completed on substrate encapsulation by GroES. (4) Folding in the chaperonin cage. (5) Substrate release on GroES dissociation. (6) Rebinding of incompletely folded states. N, native state; I, folding intermediate. (Reprinted, with permission of Macmillan Publishers Ltd., from Hartl and Hayer-Hartl 2009.) (B) The Hsp90 system. Left panel: Crystal structure of the Hsp90 dimer from S. cerevisiae with the regulator p23 bound to the amino-domain (PDB 2CG9) (Ali et al. 2006). Right panel: ATPase cycle of Hsp90. On ATP binding (1) the amino-terminal ATPase domain (ND) of Hsp90 undergoes a conformational change leading to the closure of the ATP lid. After lid closure, the first 24 amino acid residues of each Hsp90 monomer dimerize and the first β-strand and α-helix swap to associate with the ND of the other monomer (2). Furthermore, in each monomer, the ND contacts the corresponding M-domain (MD). This metastable conformation is committed for ATP hydrolysis (3). This results in a compaction of the Hsp90 dimer, in which the individual monomers twist around each other. After hydrolysis (4), the NDs dissociate and both monomers separate amino-terminally. Various cofactors regulate this cycle: Cdc37, which delivers certain kinase substrates to Hsp90, inhibits the ATPase activity, and Hop, a TPR-protein that joins Hsp70 to Hsp90, inhibits ND dimerization. Aha1 stimulates ATP hydrolysis, whereas p23 stabilizes the dimerized form of Hsp90 before ATP hydrolysis. These factors are thought to adjust the kinetic properties of the cycle to achieve certain conformational transitions in Hsp90-bound client proteins, as well as their release from Hsp90. (Modified, with permission, from Wandinger et al. 2008.)
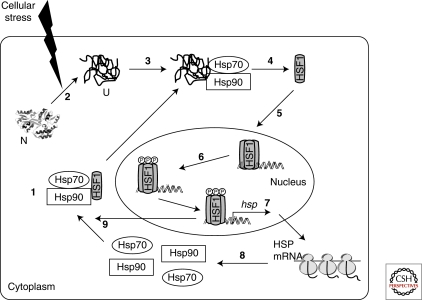
Transcriptional regulation of heat shock proteins by HSF1. (1) Under normal conditions HSF1 exists as an inert monomer in a complex with Hsp90 and Hsp70. (2) Cellular stress increases the amount of denatured proteins in the cytoplasm. (3) Denatured proteins bind to Hsp70 and Hsp90, resulting in the displacement of HSF1 (4). (5) HSF1 then trimerizes and translocates to the nucleus where it undergoes a series of posttranslational modifications, including phosphorylation (6). (7) Activated HSF1 trimer induces the transcription of a number of hsp genes, resulting in the translation of HSPs, including Hsp90 and Hsp70 (8). (9) The increased cellular concentration of these chaperones in turn inactivates HSF1 by binding to monomeric or trimeric forms of HSF1. N, native protein; U, unfolded protein. (Powers and Workman 2007.)
References
-
- Abravaya K, Myers MP, Murphy SP, Morimoto RI 1992. The human heat shock protein hsp70 interacts with HSF, the transcription factor that regulates heat shock gene expression. Genes Dev 6: 1153–1164 - PubMed
-
- Agashe VR, Guha S, Chang HC, Genevaux P, Hayer-Hartl M, Stemp M, Georgopoulos C, Hartl FU, Barral JM 2004. Function of trigger factor and DnaK in multidomain protein folding: Increase in yield at the expense of folding speed. Cell 117: 199–209 - PubMed
-
- Akerfelt M, Trouillet D, Mezger V, Sistonen L 2007. Heat shock factors at a crossroad between stress and development. Ann N Y Acad Sci 1113: 15–27 - PubMed
-
- Albanese V, Yam AYW, Baughman J, Parnot C, Frydman J 2006. Systems analyses reveal two chaperone networks with distinct functions in eukaryotic cells. Cell 124: 75–88 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources