

Emergent dynamics of fast ripples in the epileptic hippocampus
- PMID: 21123571
- PMCID: PMC6634823
- DOI: 10.1523/JNEUROSCI.3357-10.2010
Emergent dynamics of fast ripples in the epileptic hippocampus
Abstract
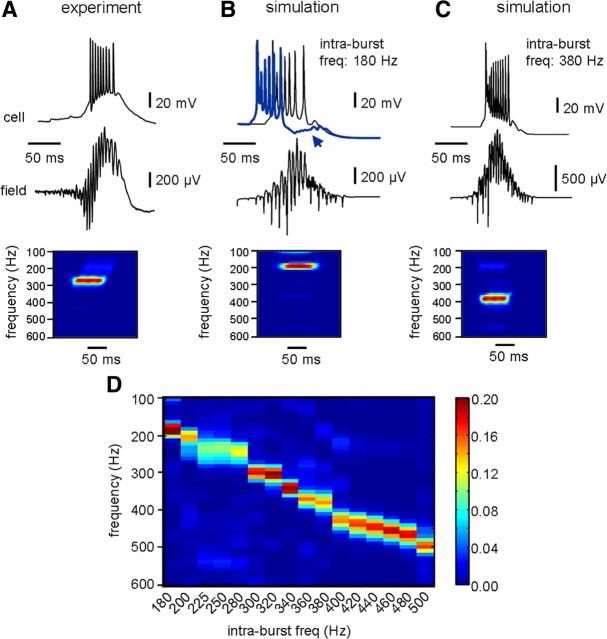
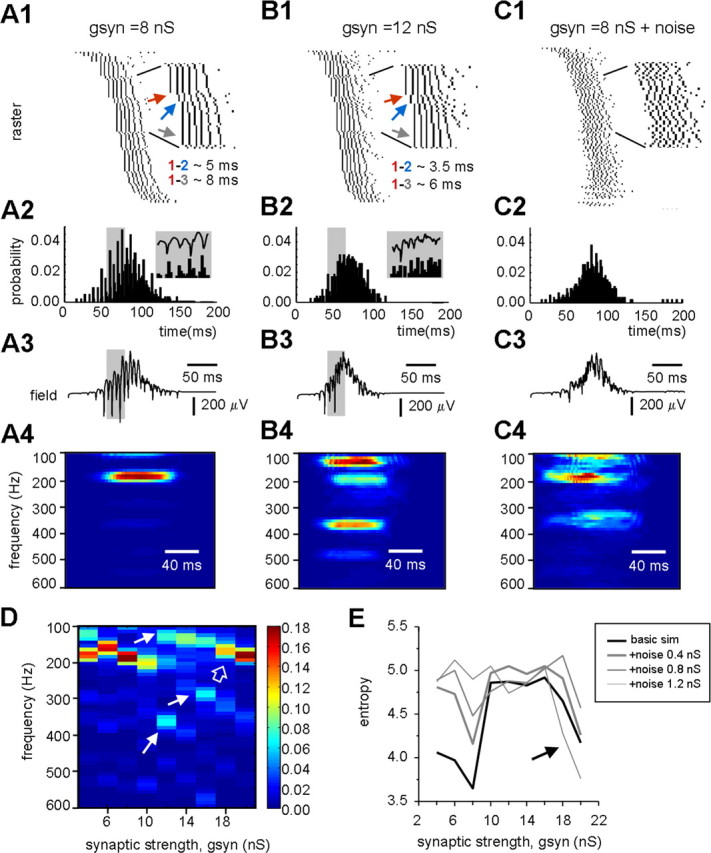
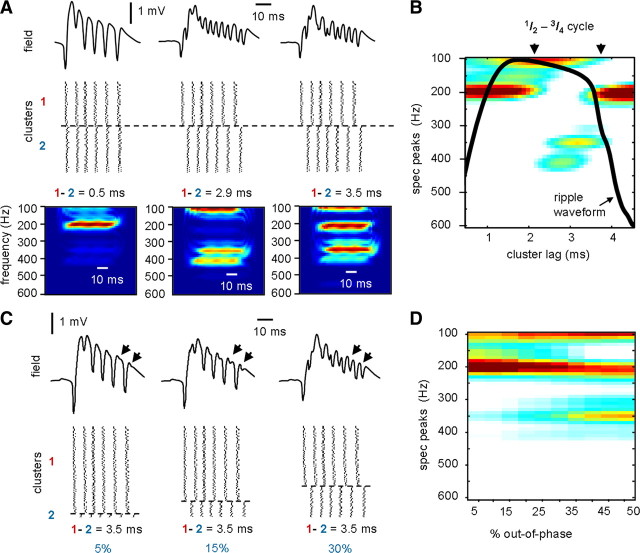
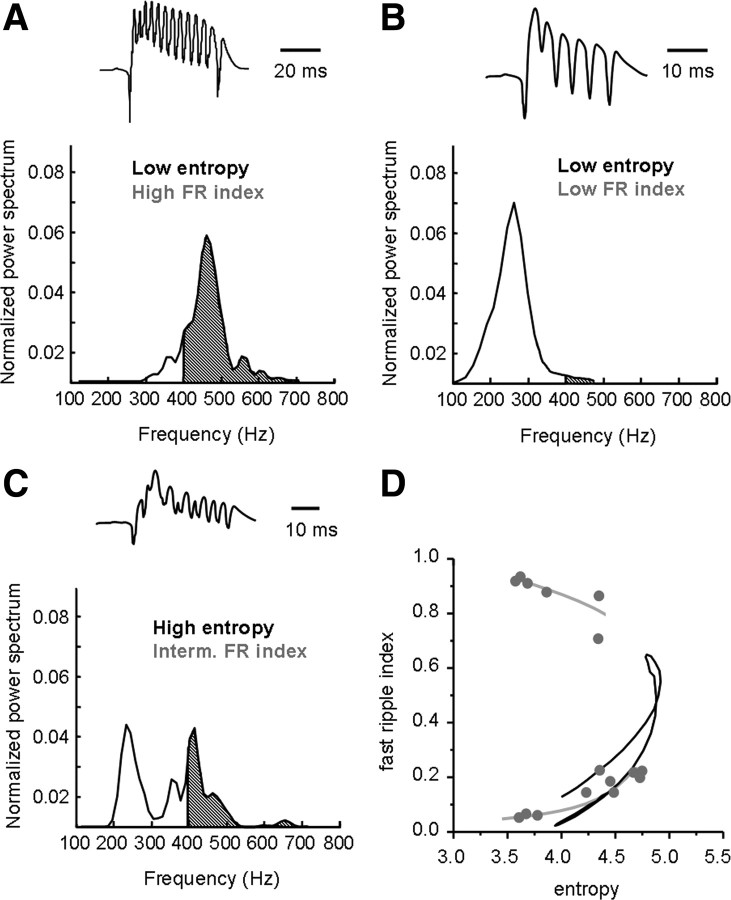
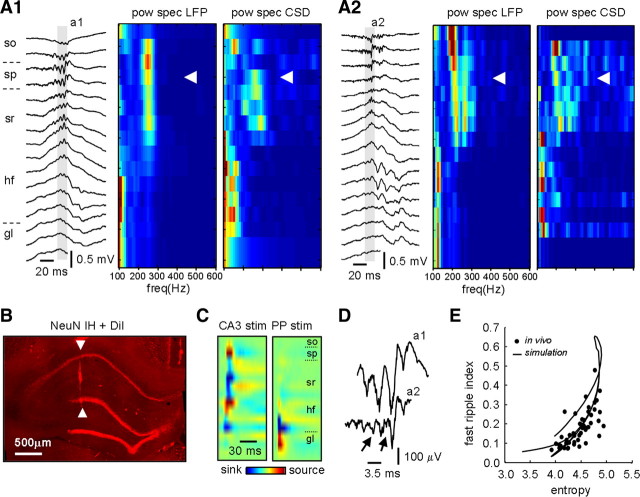
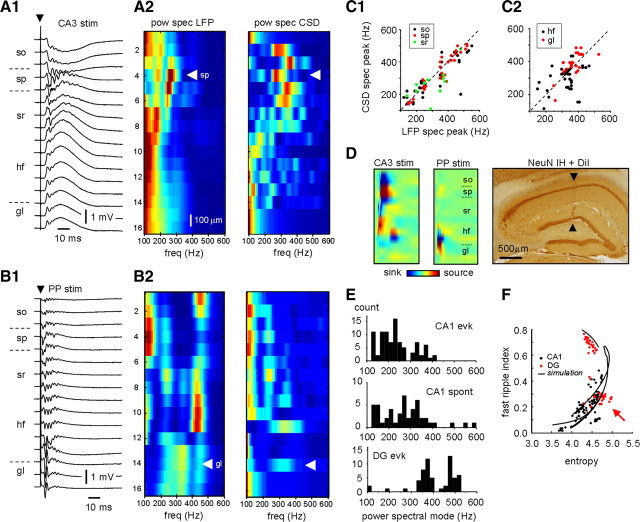
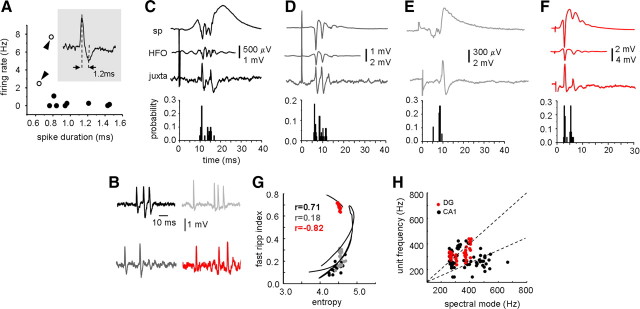
Fast ripples are a type of transient high-frequency oscillations recorded from the epileptogenic regions of the hippocampus and the temporal cortex of epileptic humans and rodents. These events presumably reflect hypersynchronous bursting of pyramidal cells. However, the oscillatory spectral content of fast ripples varies from 250 to 800 Hz, well above the maximal firing frequency of most hippocampal pyramidal neurons. How such high-frequency oscillations are generated is therefore unclear. Here, we combine computational simulations of fast ripples with multisite and juxtacellular recordings in vivo to examine the underlying mechanisms in the hippocampus of epileptic rats. We show that populations of bursting cells firing individually at 100-400 Hz can create fast ripples according to two main firing regimes: (1) in-phase synchronous firing resulting in "pure" fast ripples characterized by single spectral peaks that reflect single-cell behavior and (2) out-of-phase firing that results in "emergent" fast ripples. Using simulations, we found that fast ripples generated under these two different regimes can be quantitatively separated by their spectral characteristics, and we took advantage of this separability to examine their dynamics in vivo. We found that in-phase firing can reach frequencies up to 300 Hz in the CA1 and up to 400 Hz in the dentate gyrus. The organization of out-of-phase firing is determined by firing delays between cells discharging at low frequencies. The two firing regimes compete dynamically, alternating randomly from one fast ripple event to the next, and they reflect the functional dynamic organization of the different regions of the hippocampus.
Figures
References
-
- Adrian E. Discharge frequencies in the cerebral and cerebellar cortex. Proc Phys Soc. 1935;83:32–33.
-
- Andersen P, Bliss TVP, Skrede KK. Unit analysis of hippocampal population spikes. Exp Brain Res. 1971;13:208–221. - PubMed
-
- Bernard C, Anderson A, Becker A, Poolos NP, Beck H, Johnston D. Acquired dendritic channelopathy in temporal lobe epilepsy. Science. 2004;305:532–535. - PubMed