

Freshwater sponges have functional, sealing epithelia with high transepithelial resistance and negative transepithelial potential
- PMID: 21124779
- PMCID: PMC2993944
- DOI: 10.1371/journal.pone.0015040
Freshwater sponges have functional, sealing epithelia with high transepithelial resistance and negative transepithelial potential
Abstract
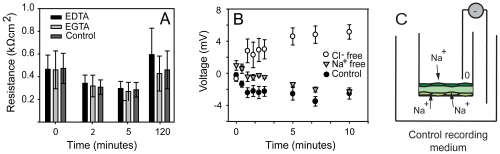
Epithelial tissue - the sealed and polarized layer of cells that regulates transport of ions and solutes between the environment and the internal milieu - is a defining characteristic of the Eumetazoa. Sponges, the most ancient metazoan phylum, are generally believed to lack true epithelia, but their ability to occlude passage of ions has never been tested. Here we show that freshwater sponges (Demospongiae, Haplosclerida) have functional epithelia with high transepithelial electrical resistance (TER), a transepithelial potential (TEP), and low permeability to small-molecule diffusion. Curiously, the Amphimedon queenslandica sponge genome lacks the classical occluding genes [5] considered necessary to regulate sealing and control of ion transport. The fact that freshwater sponge epithelia can seal suggests that either occluding molecules have been lost in some sponge lineages, or demosponges use novel molecular complexes for epithelial occlusion; if the latter, it raises the possibility that mechanisms for occlusion used by sponges may exist in other metazoa. Importantly, our results imply that functional epithelia evolved either several times, or once, in the ancestor of the Metazoa.
Conflict of interest statement
Figures



References
-
- Philippe H, Derelle R, Lopez P, Pick K, Borchiellini C, et al. Phylogenomics Revives Traditional Views on Deep Animal Relationships. Curr Biol. 2009;19:706–712. - PubMed
-
- Love GD, Grosjean E, Stalvies C, Fike DA, Grotzinger JP, et al. Fossil steroids record the appearance of Demospongiae during the Cryogenian period. Nature. 2009;457:718–721. - PubMed
-
- Mackie GO. Introduction to the diploblastic level. In: Bereiter-Hahn J, Matoltsy AG, Richards KS, editors. Biology of the integument. Berlin: Springer-Verlag; 1984. pp. 43–46.
-
- Cereijido M, Contreras RG, Shoshani L. Cell adhesion, polarity, and epithelia in the dawn of metazoans. Physiol Rev. 2004;84:1229–1262. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
