

Surface properties of Helicobacter pylori urease complex are essential for persistence
- PMID: 21124783
- PMCID: PMC2993952
- DOI: 10.1371/journal.pone.0015042
Surface properties of Helicobacter pylori urease complex are essential for persistence
Abstract
The enzymatic activity of Helicobacter pylori's urease neutralises stomach acidity, thereby promoting infection by this pathogen. Urease protein has also been found to interact with host cells in vitro, although this property's possible functional importance has not been studied in vivo. To test for a role of the urease surface in the host/pathogen interaction, surface exposed loops that display high thermal mobility were targeted for inframe insertion mutagenesis. H. pylori expressing urease with insertions at four of eight sites tested retained urease activity, which in three cases was at least as stable as was wild-type urease at pH 3. Bacteria expressing one of these four mutant ureases, however, failed to colonise mice for even two weeks, and a second had reduced bacterial titres after longer term (3 to 6 months) colonisation. These results indicate that a discrete surface of the urease complex is important for H. pylori persistence during gastric colonisation. We propose that this surface interacts directly with host components important for the host-pathogen interaction, immune modulation or other actions that underlie H. pylori persistence in its special gastric mucosal niche.
Conflict of interest statement
Figures
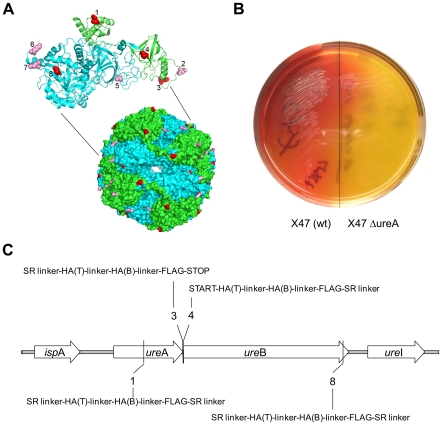
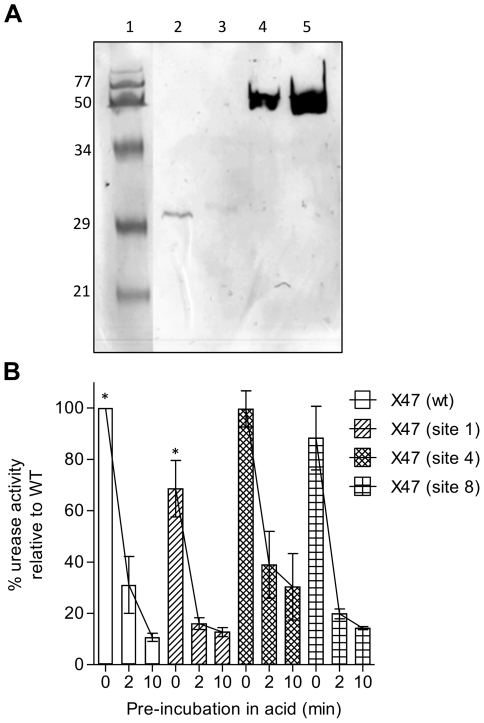
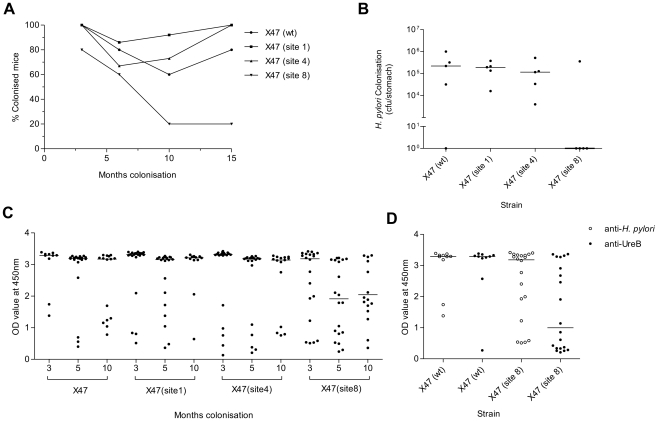
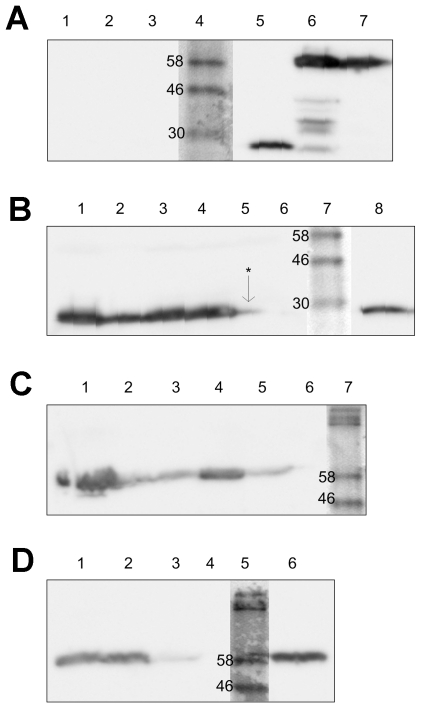

References
-
- Suzuki T, Matsuo K, Ito H, Hirose K, Wakai K, et al. A past history of gastric ulcers and Helicobacter pylori infection increase the risk of gastric malignant lymphoma. Carcinogenesis. 2006. - PubMed
-
- Marcus EA, Scott DR. Cell lysis is responsible for the appearance of extracellular urease in Helicobacter pylori. Helicobacter. 2001;6:93–99. - PubMed
-
- Williams CL, Preston T, Hossack M, Slater C, McColl KEL. Helicobacter pylori utilises urease for amino acid synthesis. FEMS Immunol Med Microbiol. 1996;13:87–94. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
