

Systematic dissection and trajectory-scanning mutagenesis of the molecular interface that ensures specificity of two-component signaling pathways
- PMID: 21124821
- PMCID: PMC2991266
- DOI: 10.1371/journal.pgen.1001220
Systematic dissection and trajectory-scanning mutagenesis of the molecular interface that ensures specificity of two-component signaling pathways
Abstract
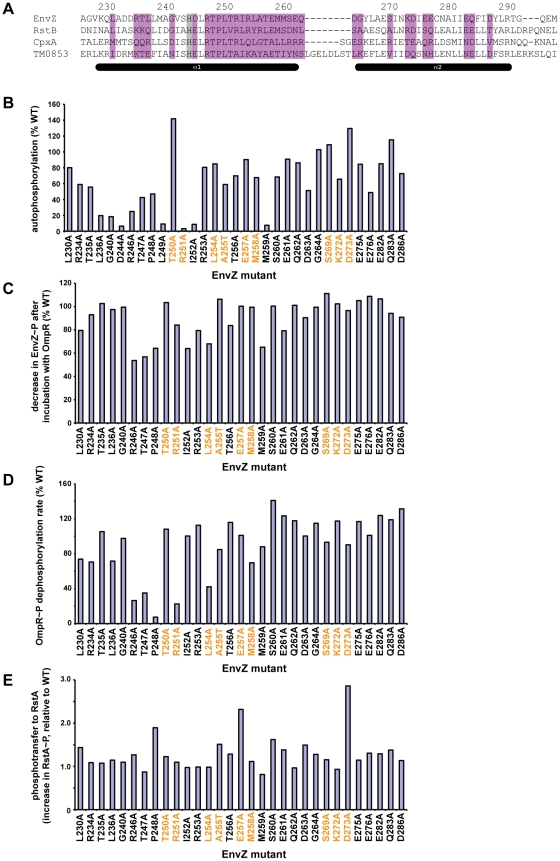
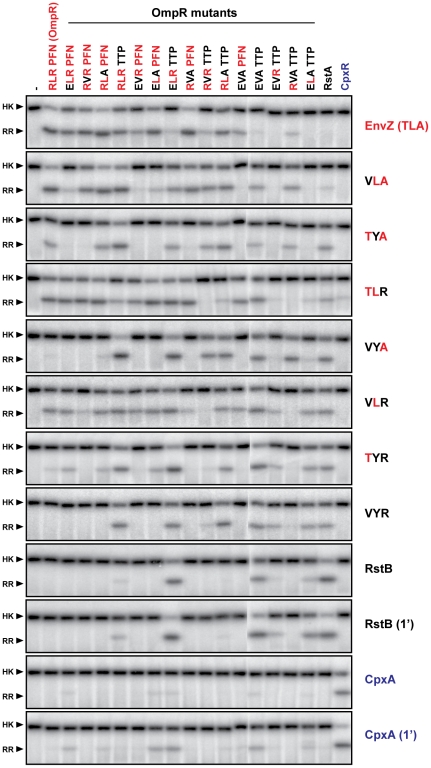
Two-component signal transduction systems enable bacteria to sense and respond to a wide range of environmental stimuli. Sensor histidine kinases transmit signals to their cognate response regulators via phosphorylation. The faithful transmission of information through two-component pathways and the avoidance of unwanted cross-talk require exquisite specificity of histidine kinase-response regulator interactions to ensure that cells mount the appropriate response to external signals. To identify putative specificity-determining residues, we have analyzed amino acid coevolution in two-component proteins and identified a set of residues that can be used to rationally rewire a model signaling pathway, EnvZ-OmpR. To explore how a relatively small set of residues can dictate partner selectivity, we combined alanine-scanning mutagenesis with an approach we call trajectory-scanning mutagenesis, in which all mutational intermediates between the specificity residues of EnvZ and another kinase, RstB, were systematically examined for phosphotransfer specificity. The same approach was used for the response regulators OmpR and RstA. Collectively, the results begin to reveal the molecular mechanism by which a small set of amino acids enables an individual kinase to discriminate amongst a large set of highly-related response regulators and vice versa. Our results also suggest that the mutational trajectories taken by two-component signaling proteins following gene or pathway duplication may be constrained and subject to differential selective pressures. Only some trajectories allow both the maintenance of phosphotransfer and the avoidance of unwanted cross-talk.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Schwartz MA, Madhani HD. Principles of MAP kinase signaling specificity in Saccharomyces cerevisiae. Annu Rev Genet. 2004;38:725–748. - PubMed
-
- Ubersax JA, Ferrell JE., Jr Mechanisms of specificity in protein phosphorylation. Nat Rev Mol Cell Biol. 2007;8:530–541. - PubMed
-
- Stock AM, Robinson VL, Goudreau PN. Two-component signal transduction. Annu Rev Biochem. 2000;69:183–215. - PubMed
-
- Laub MT, Goulian M. Specificity in two-component signal transduction pathways. Annu Rev Genet. 2007;41:121–145. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
