

Suppressive effects of vascular endothelial growth factor-B on tumor growth in a mouse model of pancreatic neuroendocrine tumorigenesis
- PMID: 21124841
- PMCID: PMC2991338
- DOI: 10.1371/journal.pone.0014109
Suppressive effects of vascular endothelial growth factor-B on tumor growth in a mouse model of pancreatic neuroendocrine tumorigenesis
Abstract
Background: The family of vascular endothelial growth factors (VEGF) contains key regulators of blood and lymph vessel development, including VEGF-A, -B, -C, -D, and placental growth factor. The role of VEGF-B during physiological or pathological angiogenesis has not yet been conclusively delineated. Herein, we investigate the function of VEGF-B by the generation of mouse models of cancer with transgenic expression of VEGF-B or homozygous deletion of Vegfb.
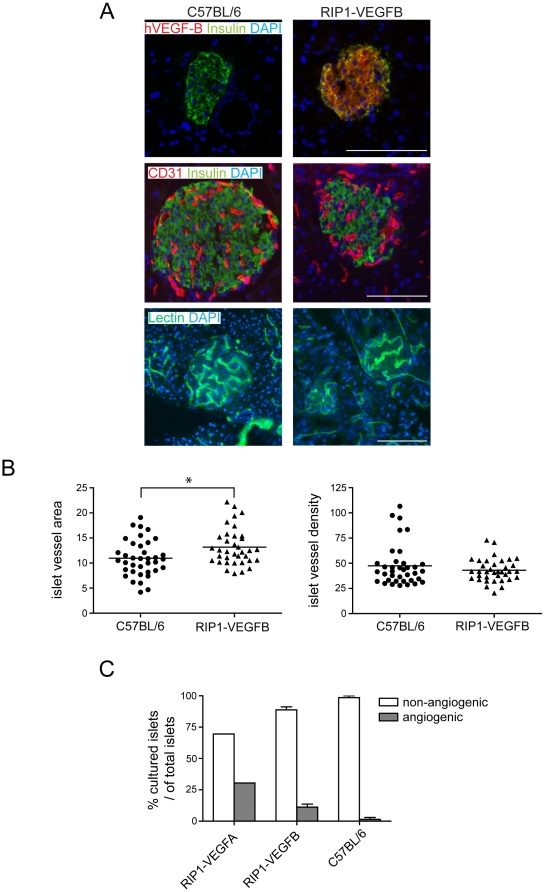
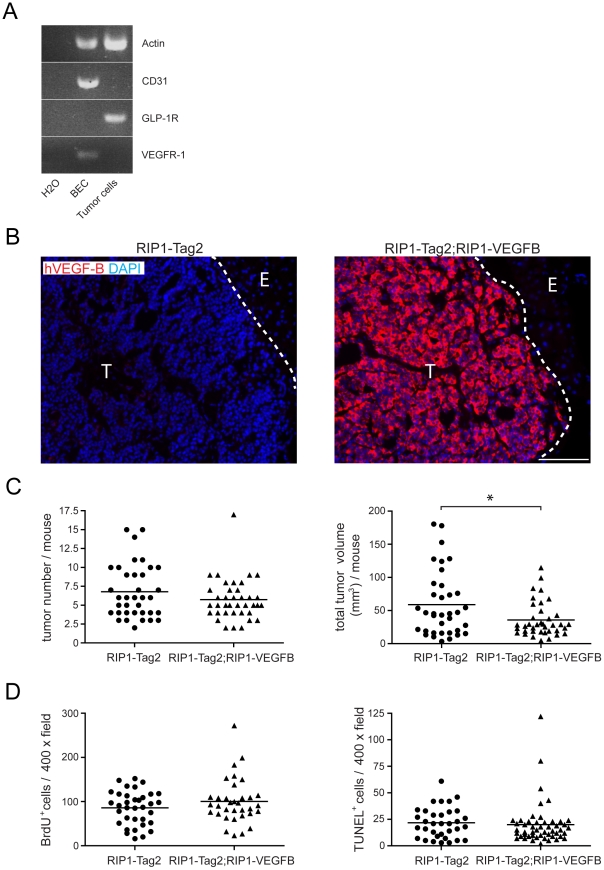
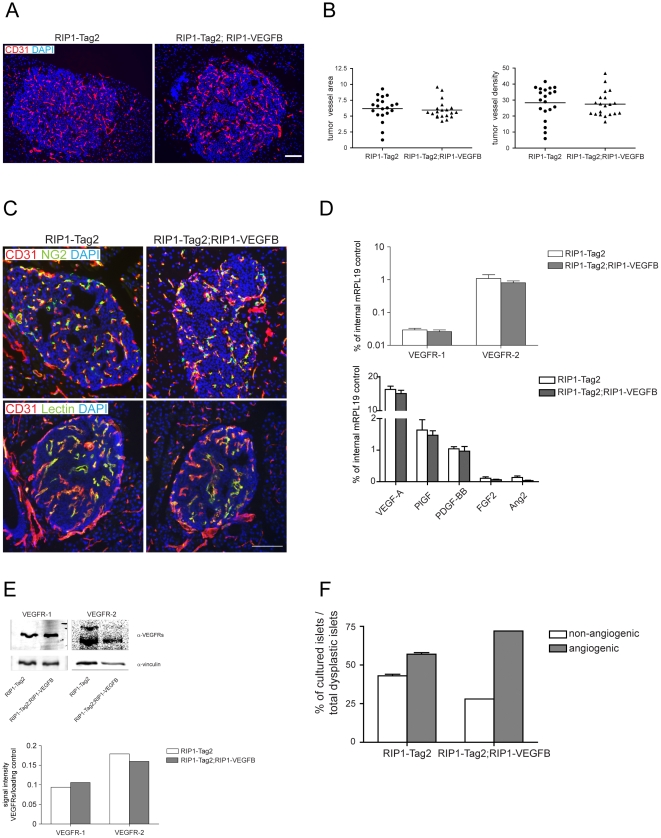
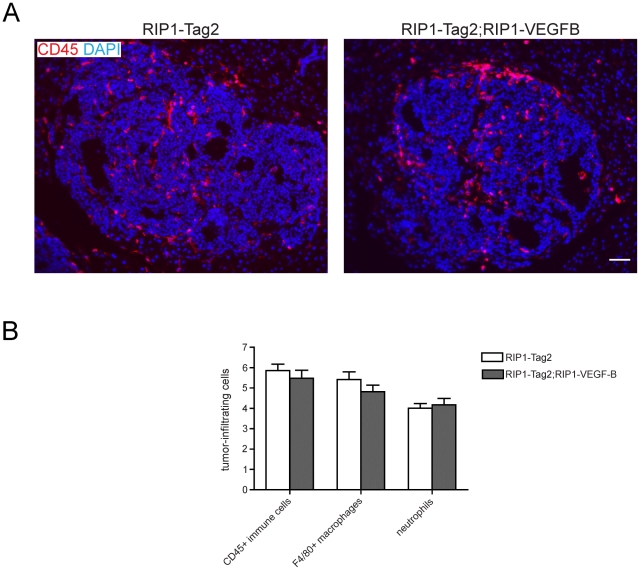
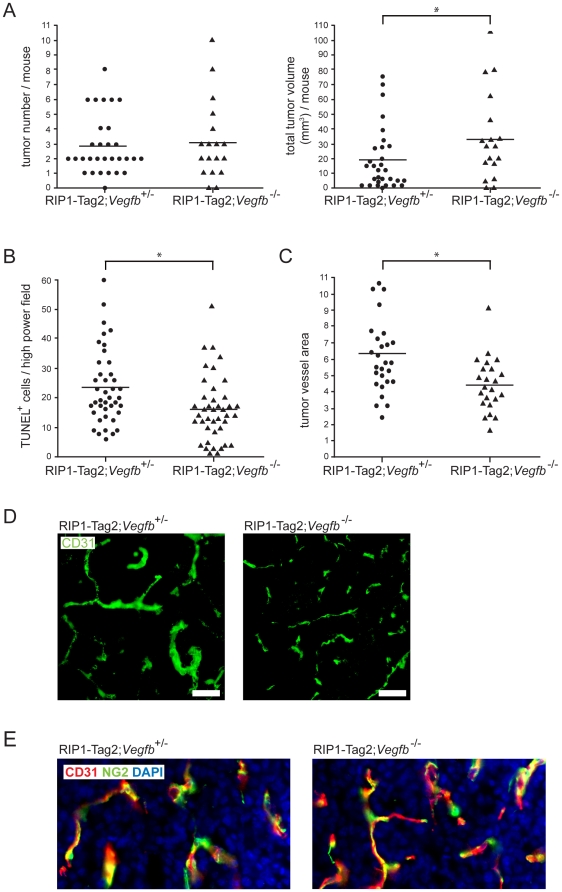
Methodology/principal findings: Ectopic expression of VEGF-B in the insulin-producing β-cells of the pancreas did not alter the abundance or architecture of the islets of Langerhans. The vasculature from transgenic mice exhibited a dilated morphology, but was of similar density as that of wildtype mice. Unexpectedly, we found that transgenic expression of VEGF-B in the RIP1-Tag2 mouse model of pancreatic neuroendocrine tumorigenesis retarded tumor growth. Conversely, RIP1-Tag2 mice deficient for Vegfb presented with larger tumors. No differences in vascular density, perfusion or immune cell infiltration upon altered Vegfb gene dosage were noted. However, VEGF-B acted to increase blood vessel diameter both in normal pancreatic islets and in RIP1-Tag2 tumors.
Conclusions/significance: Taken together, our results illustrate the differences in biological function between members of the VEGF family, and highlight the necessity of in-depth functional studies of VEGF-B to fully understand the effects of VEGFR-1 inhibitors currently used in the clinic.
Conflict of interest statement
Figures
References
-
- Carmeliet P. Angiogenesis in life, disease and medicine. Nature. 2005;438:932–936. - PubMed
-
- Olsson AK, Dimberg A, Kreuger J, Claesson-Welsh L. VEGF receptor signalling - in control of vascular function. Nat Rev Mol Cell Biol. 2006;7:359–371. - PubMed
-
- Carmeliet P, Ferreira V, Breier G, Pollefeyt S, Kieckens L, et al. Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature. 1996;380:435–439. - PubMed
-
- Ferrara N, Carver-Moore K, Chen H, Dowd M, Lu L, et al. Heterozygous embryonic lethality induced by targeted inactivation of the VEGF gene. Nature. 1996;380:439–442. - PubMed
-
- Fischer C, Jonckx B, Mazzone M, Zacchigna S, Loges S, et al. Anti-PlGF inhibits growth of VEGF(R)-inhibitor-resistant tumors without affecting healthy vessels. Cell. 2007;131:463–475. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
