

Antioxidants, redox signaling, and pathophysiology in schizophrenia: an integrative view
- PMID: 21126177
- PMCID: PMC3159108
- DOI: 10.1089/ars.2010.3603
Antioxidants, redox signaling, and pathophysiology in schizophrenia: an integrative view
Abstract
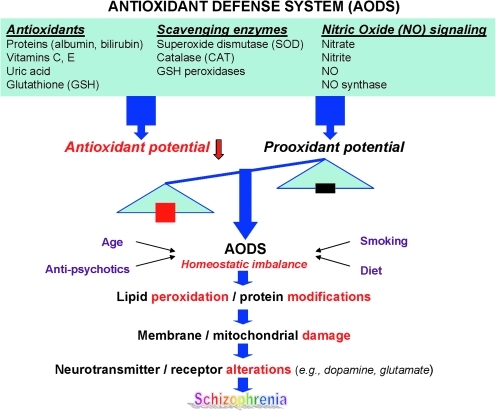
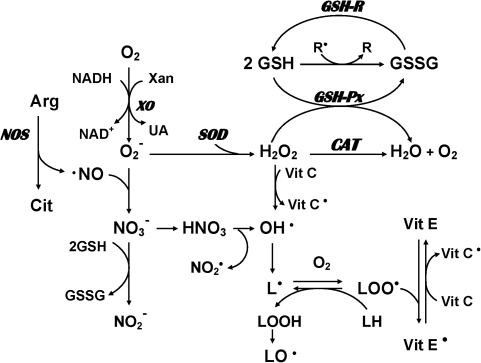
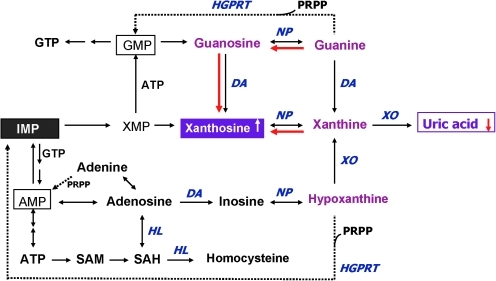
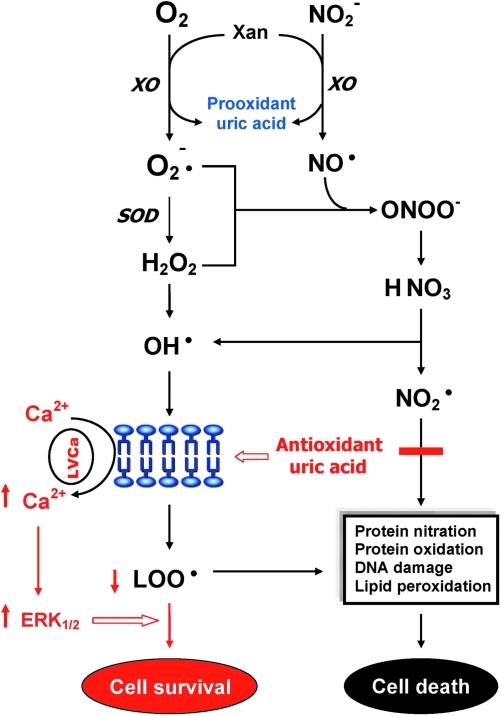
Schizophrenia (SZ) is a brain disorder that has been intensively studied for over a century; yet, its etiology and multifactorial pathophysiology remain a puzzle. However, significant advances have been made in identifying numerous abnormalities in key biochemical systems. One among these is the antioxidant defense system (AODS) and redox signaling. This review summarizes the findings to date in human studies. The evidence can be broadly clustered into three major themes: perturbations in AODS, relationships between AODS alterations and other systems (i.e., membrane structure, immune function, and neurotransmission), and clinical implications. These domains of AODS have been examined in samples from both the central nervous system and peripheral tissues. Findings in patients with SZ include decreased nonenzymatic antioxidants, increased lipid peroxides and nitric oxides, and homeostatic imbalance of purine catabolism. Reductions of plasma antioxidant capacity are seen in patients with chronic illness as well as early in the course of SZ. Notably, these data indicate that many AODS alterations are independent of treatment effects. Moreover, there is burgeoning evidence indicating a link among oxidative stress, membrane defects, immune dysfunction, and multineurotransmitter pathologies in SZ. Finally, the body of evidence reviewed herein provides a theoretical rationale for the development of novel treatment approaches.
Figures
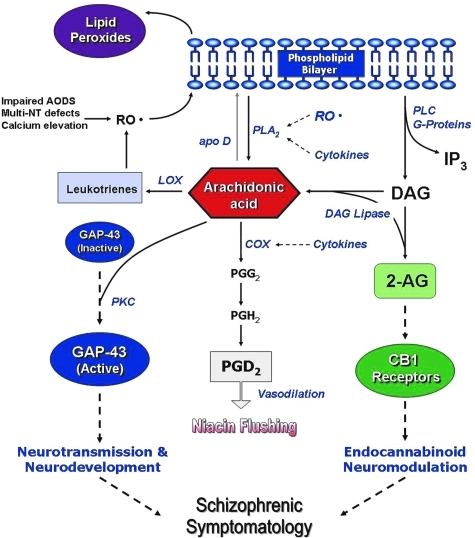
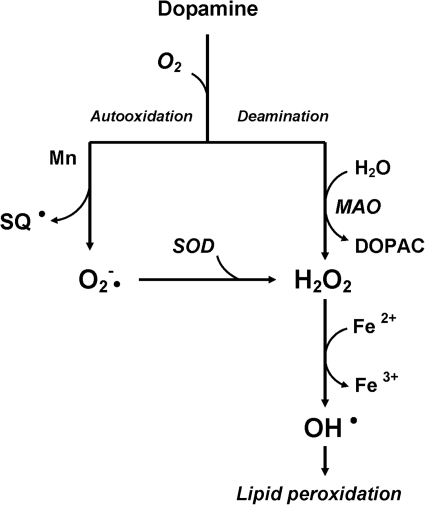
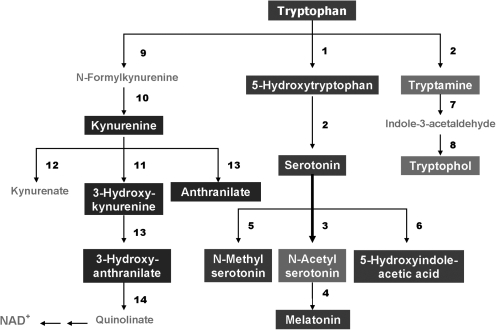
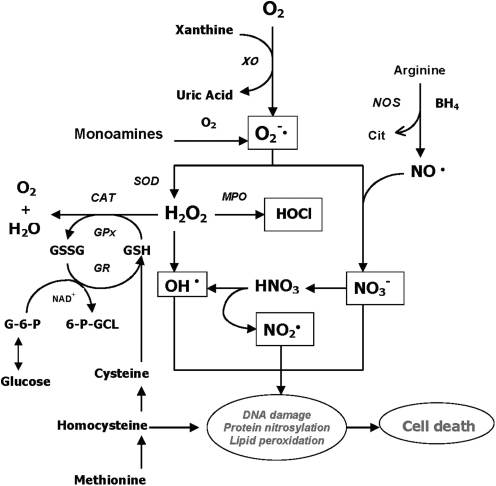
References
-
- Abdalla DS. Manteiro HP. Olivera JA. Bechara EJ. Activities of superoxide dismutase and glutathione peroxidase in schizophrenic and manic depressive patients. Clin Chem. 1986;32:805–807. - PubMed
-
- Akyol O. Zoroglu S. Armutcu F. Sahin S. Gurel A. Nitric oxide as a physiopathological factor in neuropsychiatric disorders. In Vivo. 2004;18:377–390. - PubMed
-
- Al-Chalabi BM. Thanoon IA. Ahmed FA. Potential effect of olanzapine on total antioxidant status and lipid peroxidation in schizophrenic patients. Neuropsychobiology. 2009;59:8–11. - PubMed
-
- Amminger GP. Schäfer MR. Papageorgiou K. Klier CM. Cotton SM. Harrigan SM. Mackinnon A. McGorry PD. Berger GE. Long-chain omega-3 fatty acids for indicated prevention of psychotic disorders: a randomized, placebo-controlled trial. Arch Gen Psychiatry. 2010;67:146–154. - PubMed
-
- Anderson ME. Glutathione: an overview of biosynthesis and modulation. Chem Biol Interact. 1998;111–112:1–14. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
