

A novel web-based TinT application and the chronology of the Primate Alu retroposon activity
- PMID: 21126360
- PMCID: PMC3014933
- DOI: 10.1186/1471-2148-10-376
A novel web-based TinT application and the chronology of the Primate Alu retroposon activity
Abstract
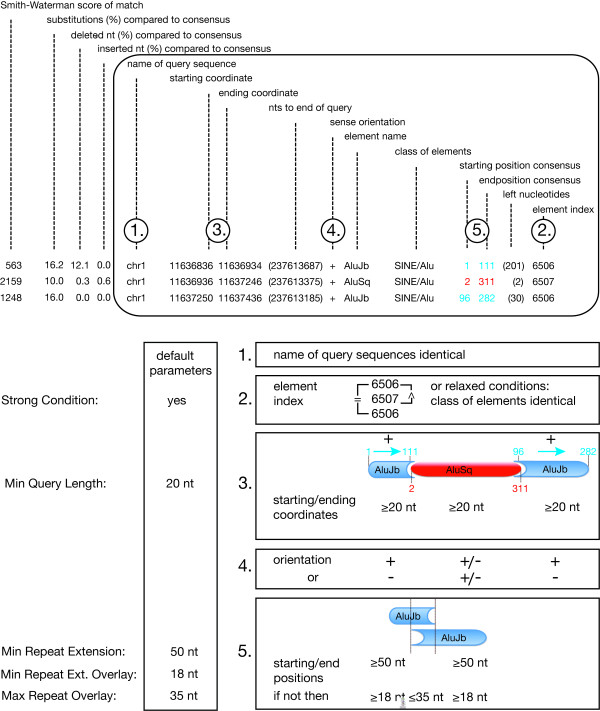
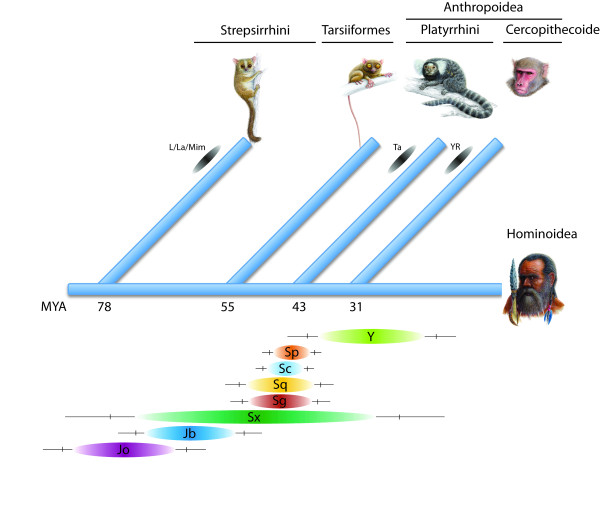
Background: DNA sequences afford access to the evolutionary pathways of life. Particularly mobile elements that constantly co-evolve in genomes encrypt recent and ancient information of their host's history. In mammals there is an extraordinarily abundant activity of mobile elements that occurs in a dynamic succession of active families, subfamilies, types, and subtypes of retroposed elements. The high frequency of retroposons in mammals implies that, by chance, such elements also insert into each other. While inactive elements are no longer able to retropose, active elements retropose by chance into other active and inactive elements. Thousands of such directional, element-in-element insertions are found in present-day genomes. To help analyze these events, we developed a computational algorithm (Transpositions in Transpositions, or TinT) that examines the different frequencies of nested transpositions and reconstructs the chronological order of retroposon activities.
Results: By examining the different frequencies of such nested transpositions, the TinT application reconstructs the chronological order of retroposon activities. We use such activity patterns as a comparative tool to (1) delineate the historical rise and fall of retroposons and their relations to each other, (2) understand the retroposon-induced complexity of recent genomes, and (3) find selective informative homoplasy-free markers of phylogeny. The efficiency of the new application is demonstrated by applying it to dimeric Alu Short INterspersed Elements (SINE) to derive a complete chronology of such elements in primates.
Conclusion: The user-friendly, web-based TinT interface presented here affords an easy, automated screening for nested transpositions from genome assemblies or trace data, assembles them in a frequency-matrix, and schematically displays their chronological activity history.
Figures

References
-
- Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, Devon K, Dewar K, Doyle M, FitzHugh W, Funke R, Gage D, Harris K, Heaford A, Howland J, Kann L, Lehoczky J, LeVine R, McEwan P, McKernan K, Meldrim J, Mesirov JP, Miranda C, Morris W, Naylor J, Raymond C, Rosetti M, Santos R, Sheridan A, Sougnez C. et al. Initial sequencing and analysis of the human genome. Nature. 2001;409(6822):860–921. doi: 10.1038/35057062. - DOI - PubMed
-
- Hillier LD, Miller W, Birney E, Warren WC, Hardison RC, Ponting CP, Bork P, Burt DW, Groenen MAM, Delany ME, Dodgson JB, Chinwalla AT, Cliften PF, Clifton SW, Delehaunty KD, Fronick C, Fulton RS, Graves TA, Kremitzki C, Layman D, Magrini V, McPherson JD, Miner TL, Minx P, Nash WE, Nhan MN, Nelson JO, Oddy LG, Pohl CS, Randall-Maher J. et al. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature. 2004;432(7018):695–716. doi: 10.1038/nature03154. - DOI - PubMed
-
- Jurka J, Klonowski P, Trifonov EN. Mammalian retroposons integrate at kinkable DNA sites. J Biomol Struct Dyn. 1998;15(4):717–721. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
