

Repeat expansion affects both transcription initiation and elongation in friedreich ataxia cells
- PMID: 21127046
- PMCID: PMC3039332
- DOI: 10.1074/jbc.M110.194035
Repeat expansion affects both transcription initiation and elongation in friedreich ataxia cells
Abstract
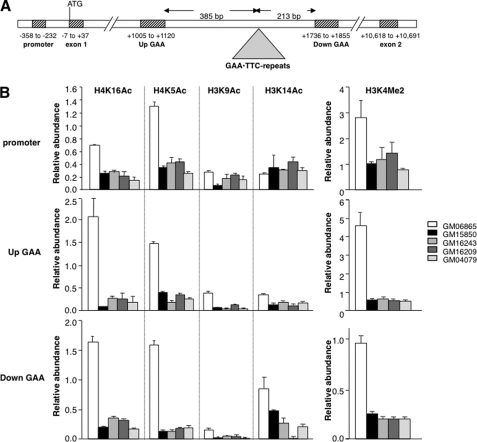
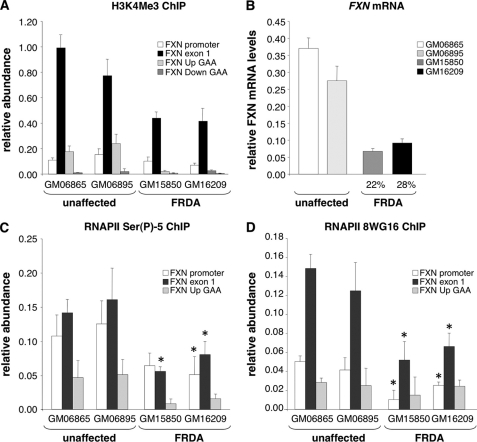
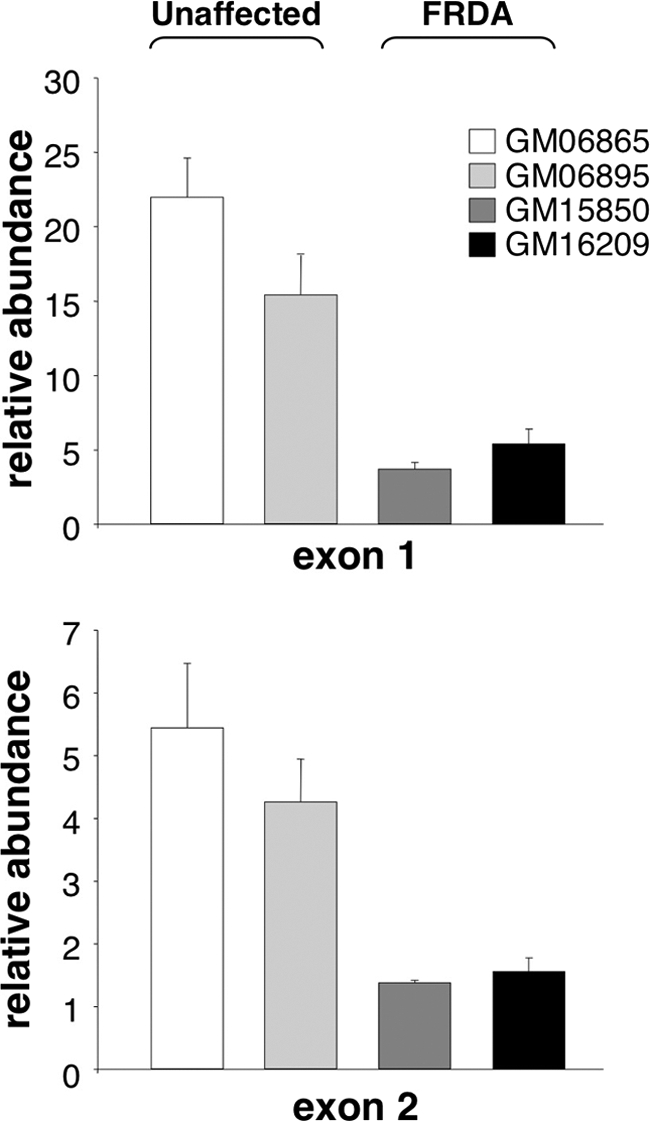
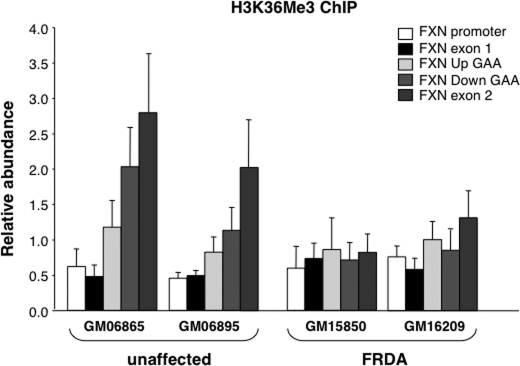
Expansion of a GAA · TTC repeat in the first intron of the frataxin (FXN) gene causes an mRNA deficit that results in Friedreich ataxia (FRDA). The region flanking the repeat on FRDA alleles is associated with more extensive DNA methylation than is seen on normal alleles and histone modifications typical of repressed genes. However, whether these changes are responsible for the mRNA deficit is controversial. Using chromatin immunoprecipitation and cell lines from affected and unaffected individuals, we show that certain marks of active chromatin are also reduced in the promoter region of the FXN gene in patient cells. Thus, the promoter chromatin may be less permissive for transcription initiation than it is on normal alleles. Furthermore, we show that the initiating form of RNA polymerase II and histone H3 trimethylated on lysine 4, a chromatin mark tightly linked to transcription initiation, are both present at lower levels on FRDA alleles. In addition, a mark of transcription elongation, trimethylated H3K36, shows a reduced rate of accumulation downstream of the repeat. Our data thus suggest that repeat expansion reduces both transcription initiation and elongation in FRDA cells. Our findings may have implications for understanding the mechanism responsible for FRDA as well as for therapeutic approaches to reverse the transcription deficit.
Figures

References
-
- Pandolfo M. (2002) Adv. Exp. Med. Biol. 516, 99–118 - PubMed
-
- Montermini L., Richter A., Morgan K., Justice C. M., Julien D., Castellotti B., Mercier J., Poirier J., Capozzoli F., Bouchard J. P., Lemieux B., Mathieu J., Vanasse M., Seni M. H., Graham G., Andermann F., Andermann E., Melançon S. B., Keats B. J., Di Donato S., Pandolfo M. (1997) Ann. Neurol. 41, 675–682 - PubMed
-
- Herman D., Jenssen K., Burnett R., Soragni E., Perlman S. L., Gottesfeld J. M. (2006) Nat. Chem. Biol. 2, 551–558 - PubMed
-
- Al-Mahdawi S., Pinto R. M., Ismail O., Varshney D., Lymperi S., Sandi C., Trabzuni D., Pook M. (2008) Hum. Mol. Genet 17, 735–746 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
