

Cytoplasmic partitioning of P granule components is not required to specify the germline in C. elegans
- PMID: 21127218
- PMCID: PMC3072820
- DOI: 10.1126/science.1193697
Cytoplasmic partitioning of P granule components is not required to specify the germline in C. elegans
Abstract
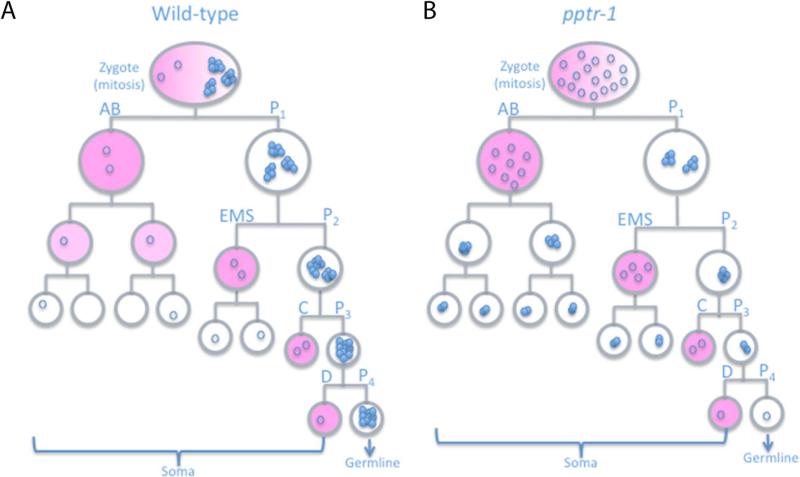
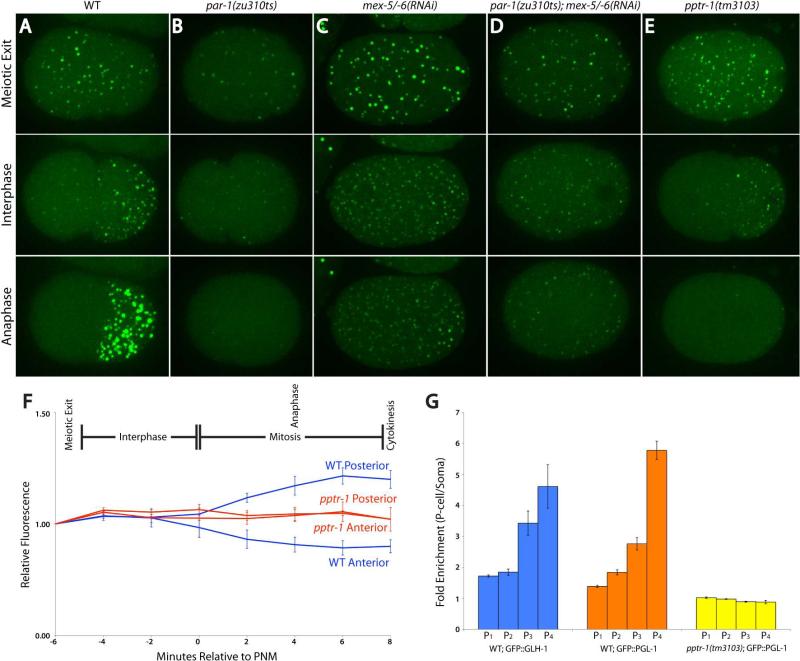
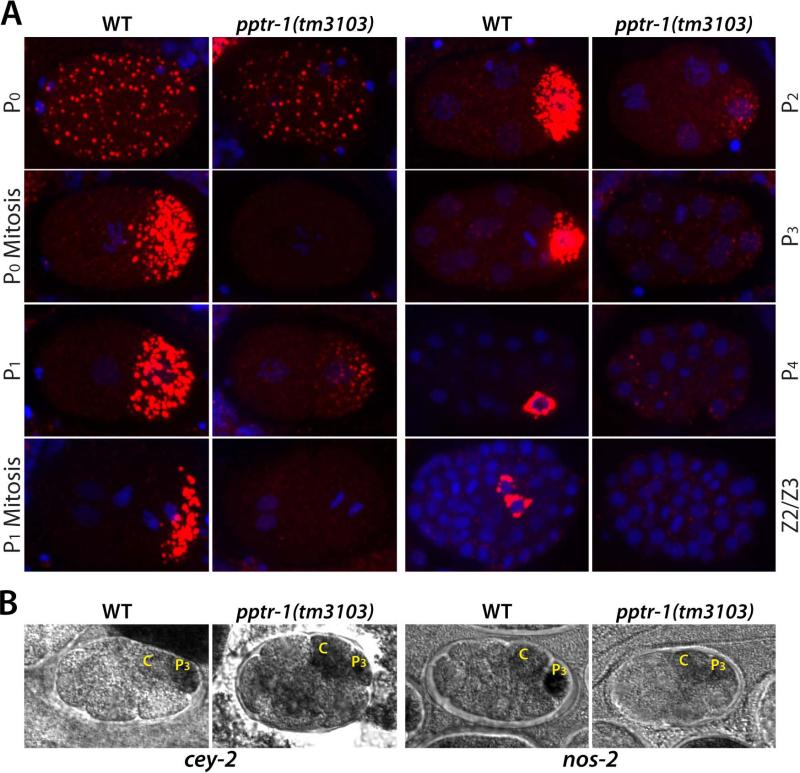
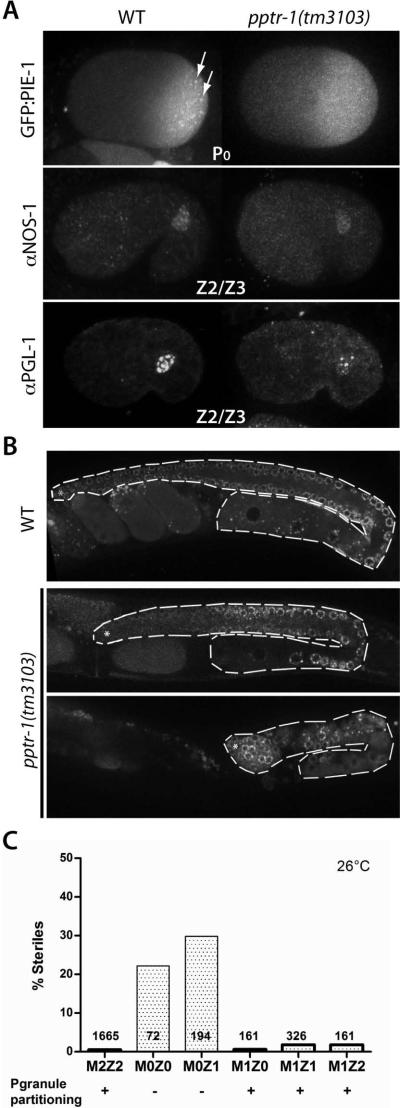
Asymmetric segregation of P granules during the first four divisions of the Caenorhabditis elegans embryo is a classic example of cytoplasmic partitioning of germline determinants. It is thought that asymmetric partitioning of P granule components during mitosis is essential to distinguish germline from soma. We have identified a mutant (pptr-1) in which P granules become unstable during mitosis and P granule proteins and RNAs are distributed equally to somatic and germline blastomeres. Despite symmetric partitioning of P granule components, pptr-1 mutants segregate a germline that uniquely expresses P granules during postembryonic development. pptr-1 mutants are fertile, except at high temperatures. Hence, asymmetric partitioning of maternal P granules is not essential to specify germ cell fate. Instead, it may serve to protect the nascent germline from stress.
Figures
Similar articles
-
MEG-1 and MEG-2 are embryo-specific P-granule components required for germline development in Caenorhabditis elegans.Genetics. 2008 Jan;178(1):295-306. doi: 10.1534/genetics.107.080218. Genetics. 2008. PMID: 18202375 Free PMC article.
-
Germline P granules are liquid droplets that localize by controlled dissolution/condensation.Science. 2009 Jun 26;324(5935):1729-32. doi: 10.1126/science.1172046. Epub 2009 May 21. Science. 2009. PMID: 19460965
-
OMA-1 is a P granules-associated protein that is required for germline specification in Caenorhabditis elegans embryos.Genes Cells. 2006 Apr;11(4):383-96. doi: 10.1111/j.1365-2443.2006.00945.x. Genes Cells. 2006. PMID: 16611242
-
Specification of the germ line.WormBook. 2005 Jul 28:1-10. doi: 10.1895/wormbook.1.9.1. WormBook. 2005. PMID: 18050414 Free PMC article. Review.
-
The P Granules of C. elegans: A Genetic Model for the Study of RNA-Protein Condensates.J Mol Biol. 2018 Nov 2;430(23):4702-4710. doi: 10.1016/j.jmb.2018.08.007. Epub 2018 Aug 8. J Mol Biol. 2018. PMID: 30096346 Free PMC article. Review.
Cited by
-
Going with the flow: insights from Caenorhabditis elegans zygote polarization.Philos Trans R Soc Lond B Biol Sci. 2020 Oct 12;375(1809):20190555. doi: 10.1098/rstb.2019.0555. Epub 2020 Aug 24. Philos Trans R Soc Lond B Biol Sci. 2020. PMID: 32829680 Free PMC article. Review.
-
Coupling between cytoplasmic concentration gradients through local control of protein mobility in the Caenorhabditis elegans zygote.Mol Biol Cell. 2015 Sep 1;26(17):2963-70. doi: 10.1091/mbc.E15-05-0302. Epub 2015 Jul 8. Mol Biol Cell. 2015. PMID: 26157168 Free PMC article.
-
PLK-1 regulates MEX-1 polarization in the C. elegans zygote.bioRxiv [Preprint]. 2024 Jul 26:2024.07.26.605193. doi: 10.1101/2024.07.26.605193. bioRxiv. 2024. PMID: 39091813 Free PMC article. Preprint.
-
Diverse functions of mRNA metabolism factors in stress defense and aging of Caenorhabditis elegans.PLoS One. 2014 Jul 25;9(7):e103365. doi: 10.1371/journal.pone.0103365. eCollection 2014. PLoS One. 2014. PMID: 25061667 Free PMC article.
-
Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels.Cell. 2012 May 11;149(4):753-67. doi: 10.1016/j.cell.2012.04.017. Cell. 2012. PMID: 22579281 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
