

Video force microscopy reveals the mechanics of ventral furrow invagination in Drosophila
- PMID: 21127270
- PMCID: PMC3009801
- DOI: 10.1073/pnas.1006591107
Video force microscopy reveals the mechanics of ventral furrow invagination in Drosophila
Abstract
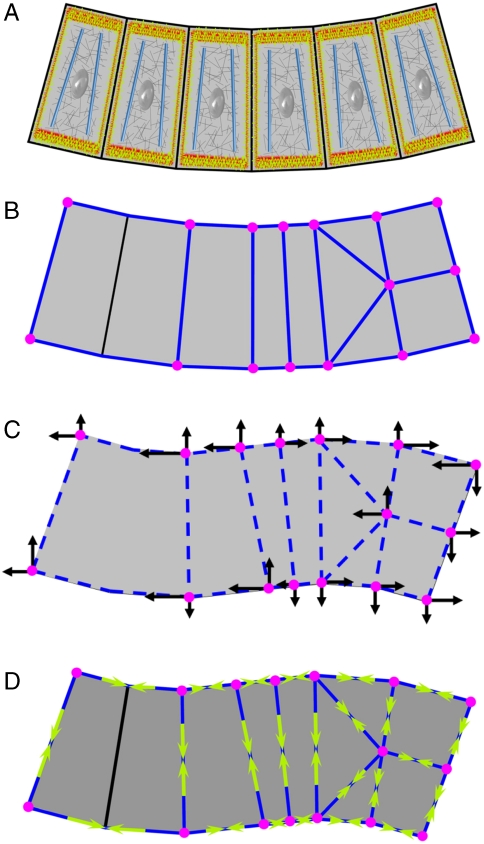
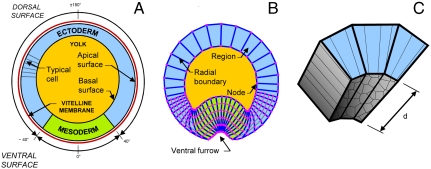
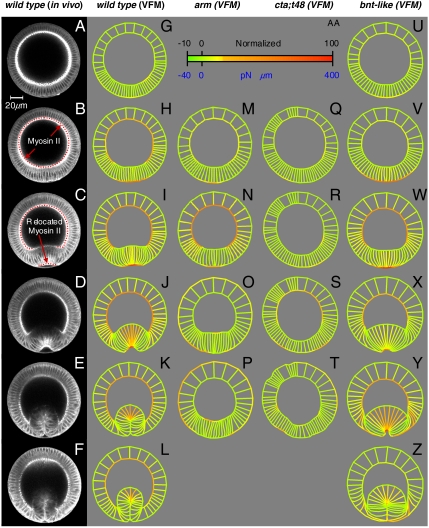
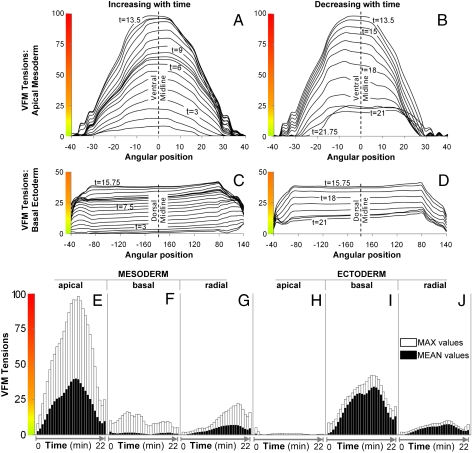
The absence of tools for mapping the forces that drive morphogenetic movements in embryos has impeded our understanding of animal development. Here we describe a unique approach, video force microscopy (VFM), that allows detailed, dynamic force maps to be produced from time-lapse images. The forces at work in an embryo are considered to be decomposed into active and passive elements, where active forces originate from contributions (e.g., actomyosin contraction) that do mechanical work to the system and passive ones (e.g., viscous cytoplasm) that dissipate energy. In the present analysis, the effects of all passive components are considered to be subsumed by an effective cytoplasmic viscosity, and the driving forces are resolved into equivalent forces along the edges of the polygonal boundaries into which the region of interest is divided. Advanced mathematical inverse methods are used to determine these driving forces. When applied to multiphoton sections of wild-type and mutant Drosophila melanogaster embryos, VFM is able to calculate the equivalent driving forces acting along individual cell edges and to do so with subminute temporal resolution. In the wild type, forces along the apical surface of the presumptive mesoderm are found to be large and to vary parabolically with time and angular position, whereas forces along the basal surface of the ectoderm, for example, are found to be smaller and nearly uniform with position. VFM shows that in mutants with reduced junction integrity and myosin II activity, the driving forces are reduced, thus accounting for ventral furrow failure.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Puech P, et al. Measuring cell adhesion forces of primary gastrulating cells from zebrafish using atomic force microscopy. J Cell Sci. 2005;118:4199–4206. - PubMed
-
- Krieg M, et al. Tensile forces govern germ-layer organization in zebrafish. Nat Cell Biol. 2008;10:429–436. - PubMed
-
- Conte V, Muñoz JJ, Baum B, Miodownik M. Robust mechanisms of ventral furrow invagination require the combination of cellular shape changes. Phys Biol. 2009;6:016010,. 10.1088/1478-3975/6/1/016010. - PubMed
-
- Chen X, Brodland GW. Multi-scale finite element modeling allows the mechanics of amphibian neurulation to be elucidated. Phys Biol. 2008;5:015003. 10.1088/1478-3975/5/1/015003. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
